



INTRODUCTION
The Yellowstone Plateau Volcanic Field hosts one of the largest and most diverse continental hydrothermal systems on Earth (Hurwitz and Lowenstern, Reference Hurwitz and Lowenstern2014). More than 10,000 thermal features are found in the volcanic field, concentrated within the Yellowstone caldera (http://rcn.montana.edu/Features/Search.aspx, accessed July 2020). Thermal areas occupy more than 48 km2 in Yellowstone National Park (Vaughan et al., Reference Vaughan, Heasler, Jaworowski, Lowenstern and Keszthelyi2014) and present-day thermal areas have been active since at least late glacial time (Muffler et al., Reference Muffler, White and Truesdell1971, Reference Muffler, White, Truesdell and Fournier1982; Fournier, Reference Fournier1989; Sturchio et al., Reference Sturchio, Pierce, Murrell and Sorey1994; Pierce et al., Reference Pierce, Cannon, Meyer, Trebesch, Watts and Morgan2007a).
The influence of hydrothermal activity on Yellowstone's terrestrial and limnologic ecosystems is poorly known but seems to occur on many time scales. At present, for example, shifts in the flow of hot, mineral-saturated fluids lead to plant mortality (Channing and Edwards, Reference Channing and Edwards2003), and heated soils change vegetation communities and stunt tree growth (Miller, Reference Miller1968; Pitt and Hutchinson, Reference Pitt and Hutchinson1982; White et al., Reference White, Hutchinson and Keith1988; Lowenstern et al., Reference Lowenstern, Heasler and Smith2003). On decadal time scales, a newer study finds that a persistent drought during the Medieval Climate Anomaly led to a shutdown of Old Faithful eruptions for several decades in the early thirteenth to middle fourteenth century, allowing conifers to encroach on the geyser mound itself (Hurwitz et al., Reference Hurwitz, King, Pederson, Martin, Damby, Manga, Hungerford and Peek2020). In postglacial time, paleoecological data suggest rapid but temporary damage to past vegetation from large hydrothermal explosions in Yellowstone Lake (Schiller et al., Reference Schiller, Whitlock, Alt and Morgan2020), and both short-lived and persistent effects, potentially driven by transformed benthic substrate, to diatom communities (Brown, Reference Brown2019). Despite these examples, we have little understanding of the long-term ecological consequences of shifting hydrothermal activity in an active thermal basin, nor are we able to evaluate the relative importance of geology and climate in shaping the ecological history of these settings.
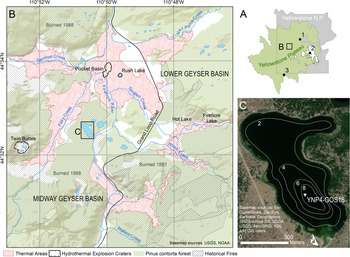
Here, we focus on the geo-ecological history of Lower Geyser Basin (LGB), the largest thermal area in Yellowstone National Park (topographic area ~25 km2, active thermal area ~14 km2) (Fig. 1B; Vaughan et al., Reference Vaughan, Heasler, Jaworowski, Lowenstern and Keszthelyi2014). A series of small lakes lies within LGB in close proximity to active thermal features. This juxtaposition enables use of lake sediment records to examine the interaction between past hydrothermal activity, climate, and ecosystem responses. An 808.5 cm-long sediment core from the largest of the LGB lakes, Goose Lake (44.54°N, 110.84°W, 2196 m asl, 13.8 ha, Fig. 1C), was studied to address the following questions: (1) What is the history of hydrothermal activity at and near Goose Lake as registered in the lake sediment archive? (2) What was the role of climate, hydrothermal activity, and fire in shaping the LGB geo-ecosystem, including its Holocene vegetation and limnologic history relative to other parts of the Yellowstone Plateau?

Figure 1. (A) Location of Lower Geyser Basin, Yellowstone National Park, Wyoming, USA, with location of referenced sites (1) Cygnet Lake, (2) Yellowstone Lake, and (3) Loon Lake relative to Yellowstone National Park and the Yellowstone Plateau (an EPA Level IV Ecoregion; Chapman et al., Reference Chapman, Bryce, Omernik, Despain, ZumBerge and Conrad2004). (B) Goose Lake location within Lower Geyser Basin, and Midway Geyser Basin, with active thermal areas indicated by red stippling (Vaughan et al., Reference Vaughan, Heasler, Jaworowski, Lowenstern and Keszthelyi2014), large hydrothermal explosion craters indicated by black ticked outlines (Morgan et al., Reference Morgan, Shanks and Pierce2009), pre-AD 1988 extent of Pinus contorta-dominated forest in green (Despain, Reference Despain1990), and extent of historical fires indicated by black hashed lines (Spatial Analysis Center, Yellowstone National Park, 2020). Evidence of hydrothermal activity is pervasive in the Goose Lake vicinity although the lake is not adjacent to any active thermal features. (C) Location of YNP4-GOS18 cores taken in the deepest part of Goose Lake. Bathymetry (m depth) modified from Arnold and Sharpe (Reference Arnold and Sharpe1967).
SITE DESCRIPTION
Situated within the 630 ka Yellowstone caldera, Lower Geyser Basin (LGB) is a large, predominantly treeless basin (Fig. 1B), surrounded by post-caldera rhyolite flows (Christiansen and Blank, Reference Christiansen and Blank1974). Within LGB, rhyolite is overlain by thick Quaternary sediment of glacial and hydrothermal provenance (Bargar and Beeson, Reference Bargar and Beeson1981; Muffler et al., Reference Muffler, White, Truesdell and Fournier1982). The area was covered by the Greater Yellowstone Glacial System during the Pinedale glaciation (locally 22–13 ka, Licciardi and Pierce, Reference Licciardi and Pierce2018). Although the timing of ice retreat in the vicinity of LGB is poorly constrained by absolute ages, the basin was probably ice-free after ~14 ka based on correlation with recessional moraines in northern Yellowstone (Licciardi and Pierce, Reference Licciardi and Pierce2008) and the basal ages of nearby lake sediment cores (Whitlock, Reference Whitlock1993). The basin's thermal features have chiefly neutral and alkaline chemistry, with fewer acidic features (http://rcn.montana.edu/Features/Search.aspx, accessed July 2020). Hydrothermal activity has occurred in LGB since at least late Pinedale time, as evidenced by thick hydrothermal deposits interbedded with glacial outwash in drilled cores (Bargar and Beeson, Reference Bargar and Beeson1981), hydrothermal explosion deposits containing proglacial lake sediments (Muffler et al., Reference Muffler, White and Truesdell1971), and silica-cemented kame deposits at several localities (Muffler et al., Reference Muffler, White, Truesdell and Fournier1982). Several large (>100 m diameter) hydrothermal explosion craters are known from LGB as well, including Twin Buttes, Pocket Basin, and Rush Lake (Muffler et al., Reference Muffler, White and Truesdell1971; Morgan et al., Reference Morgan, Shanks and Pierce2009). Evidence of present and past hydrothermal activity is pervasive throughout the Goose Lake area, including vast flats of silica sinter and diatom ooze (Muffler et al., Reference Muffler, White, Truesdell and Fournier1982), but the lake itself is ~280 m from the nearest active thermal features along Fairy Creek to the west (Fig. 1B).
LGB lies within the Yellowstone Plateau, a U.S. Environmental Protection Agency (EPA) Level IV Ecoregion (Chapman et al., Reference Chapman, Bryce, Omernik, Despain, ZumBerge and Conrad2004) that occupies the central and southwestern portions of Yellowstone National Park as well as adjacent public and private lands to the west and south, and is characterized by rhyolitic substrates and homogeneous forests of Pinus contorta (Fig. 1A). Rhyolitic lava flows, such as those surrounding LGB, weather to nutrient-poor, well-drained soils that support P. contorta to the near-exclusion of other conifers (Despain, Reference Despain1990). Within LGB, thermal activity partitions vegetation into zones based on soil temperature and, to a lesser extent, acidity (Miller, Reference Miller1968; Sheppard, Reference Sheppard1971; Rodman et al., Reference Rodman, Shovic and Thoma1996). Dense P. contorta forests grow where hydrothermal activity is not present and soils are cool (< 8°C, Rodman et al., Reference Rodman, Shovic and Thoma1996; < 12°C, Miller, Reference Miller1968) as well as patches of Artemisia tridentata and A. tripartita steppe (Anderson, H.M., personal communication, 2021). Other cool sites underlain by glacial substrate support mixed meadow with mixed grasses and herbs (e.g., Trifolium repens, Rumex acetosella, Fragaria spp., Prunella vulgaris, Chamerion spp., various Asteraceae; Sheppard, Reference Sheppard1971). Unique grassland communities grow in active thermal flats where soil temperatures reach 30–50°C and are dominated by grasses such as Puccinellia nuttalliana, Dichanthelium lanuginosum, and Danthonia spicata that remain diminutive but may grow year-round due to heated surface conditions (Sheppard, Reference Sheppard1971; Stout and Al-Niemi, Reference Stout and Al-Niemi2002). At the highest soil temperatures (>50°C), grasses are absent and the vegetation mostly consists of mosses (Sheppard, Reference Sheppard1971). LGB was extensively burned in AD 1988 and AD 1881, including around the margins of Goose Lake (Spatial Analysis Center, Yellowstone National Park, 2020).
Goose Lake (13.8 ha) has a small inlet stream to the southwest (stream flow < 0.3 m3/s; |Arnold and Sharpe, Reference Arnold and Sharpe1967) and no surface outlet. Lake bathymetry reveals a broad, shallow bench, comprising up to 80% of the lake bottom, and a maximum depth of 9.5 m (Fig. 1C; Arnold and Sharpe, Reference Arnold and Sharpe1967). The lake's origins are unclear. It may have formed from past hydrothermal processes or in a late Pinedale kettle hole, given that it is surrounded by sandy gravel deposits (Waldrop and Pierce, Reference Waldrop and Pierce1975) interpreted as glacial outwash and kames (Muffler et al., Reference Muffler, White, Truesdell and Fournier1982). Other small lakes within a 2 km radius of Goose Lake also appear to be buttressed by kame deposits (Muffler et al., Reference Muffler, White, Truesdell and Fournier1982). A recent rise in lake level is evidenced by the presence of dead, submerged pine trees along the shore. Typha latifolia, Scirpus, and Carex grow around the margin of Goose Lake, and Nuphar luteum, Potomogeton spp., Myriophyllum spp., and Persicaria amphibia are present as submerged aquatics. In an earlier study by Arnold and Sharpe (Reference Arnold and Sharpe1967), water temperatures were measured and ranged from 20.8°C at the surface to 11.6°C at the base of the hypolimnion in July, and pH was 8.1. Previous paleoecological investigations of Goose Lake include an examination of recent diatom assemblages as they relate to fish stocking (Shaw Chraïbi, Reference Shaw Chraïbi2016) and an analysis of charcoal abundance in the surface sediments following the 1988 fires (Whitlock and Millspaugh, Reference Whitlock and Millspaugh1996).
METHODS
Field sampling
A series of sediment cores (YNP4-GOS18) was collected from an anchored platform in the deepest part of Goose Lake (44.54135°N, 110.84237°W, 8.86 m water depth; Fig. 1C) in June 2018, using a modified Livingstone sampler (Wright et al., Reference Wright, Mann and Glaser1984). Cores 1A (564 cm long), 1B (100 cm long), and 1D (793 cm long) were extruded, described, and wrapped before being transported from the field. The uppermost sediments were recovered using a gravity sampler in order to retrieve the sediment-water interface, and this short core (20 cm long) was extruded in the field at 1 cm resolution and placed in Whirl-Pak® bags before transport to the lab.
Laboratory analyses
Chronology
Age control was provided by accelerator mass spectrometry (AMS) radiocarbon dating of 22 charcoal samples. Prior to submission, samples were washed with distilled water, cleaned of extraneous material with a dissecting needle under a stereomicroscope, and pretreated with acid-base-acid washes. Samples were not large enough to provide a split for δ13C analysis. Additional age control also was provided by the age of the sediment-water interface and from tephrochronology.
Lithology and geochemistry
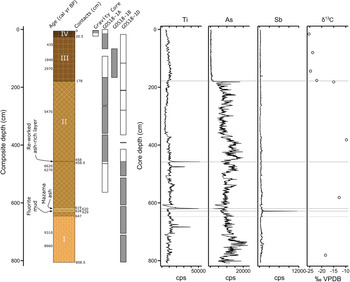
The gravity core and cores 1A, 1B, and 1D were correlated based on depth and lithostratigraphy, particularly using a distinctive, dark, ash-rich siliciclastic layer (Fig. 2). The composite core was 808.5 cm long, and subsequent descriptions use the depths of the composite core. Cores were longitudinally split, imaged, and measured every 0.5 cm for physical parameters (magnetic susceptibility, color, reflectance, density, profile) at the National Lacustrine Core Facility (LacCore, University of Minnesota-Twin Cities), and kept refrigerated except when being sampled or analyzed. Sediments were classified by composition and texture according to the scheme of Schnurrenberger et al. (Reference Schnurrenberger, Russell and Kelts2003). Measurements were made every 0.5 cm for bulk geochemistry and X-radiographs produced from scanned X-ray fluorescence (XRF) at the Large Lakes Observatory (LLO, University of Minnesota-Duluth). A section of the core with unusually high levels of antimony (Fig. 3) also was analyzed for grain texture using a scanning electron microscope (SEM), elemental composition using SEM-energy-dispersive X-ray spectroscopy (SEM-EDS), and mineralogy using X-ray diffraction (XRD) to help ascertain hydrothermal provenance at the Image and Chemical Analysis Laboratory (ICAL, Montana State University). In addition, δ13C determinations were done on bulk sediments at the U.S. Geological Survey Stable Isotope Laboratory (Reston, VA) to help ascertain sedimentary carbon provenance.

Figure 2. Lithology and geochemistry of the composite Goose Lake core YNP4-GOS18. The core was divided into four stratigraphic units: (I) Indistinctly laminated diatomaceous ooze with clay; (II) Laminated diatomaceous ooze with sapropel and minor, fragmental plant remains, punctuated by antimony-rich fluorite mud (629–628 cm depth), the Mazama ash (620–619 cm depth), and a reworked ash-rich siliciclastic deposit (458.5–458 cm depth); (III) Thinly bedded diatom ooze with sapropel; (IV) Massive diatomaceous sapropel with high water content. The diagram of the composite core is based on core depth, except the correlation between GOS18-1A and GOS18-1D, which was made using the reworked Yellowstone ash. Shaded core drives (gray) were used in developing the composite record. Abundant arsenic is associated with geothermal water influx; and antimony-rich fluorite mud (619–618 cm depth) is definitively of hydrothermal provenance. Higher δ13Csediment (‰ VPDB) likely indicates an admixture with hydrothermally degassed carbon.

Figure 3. Image of YNP4-GOS18 core section and antimony abundance from scanning XRF analysis including fluorite mud occurring at 628–629 cm depth in the composite core. (A) SEM image of sediment immediately above contact with fluorite mud; note abundant diatom frustules, sponge spicules, and fine-grained clays and organics; (B) SEM image of fluorite mud with 81 wt. % fluorite. Note large diatoms in a matrix of fluorite spherules.
Pollen analysis
Samples of 0.5 cm3 were prepared for pollen analysis at intervals of 3 cm to 16.5 cm depth in the core, with the highest sampling resolution in close proximity to ash deposits to explore potential effects of ash falls on vegetation history. Preparation followed standard procedures as outlined in Bennett and Willis (Reference Bennett, Willis, Smol, Birks, Last, Bradley and Alverson2001). A known concentration of Lycopodium spores was added as a spike to calculate pollen concentration (grains/cm3) and influx (grains/cm2/yr). Residues were preserved and mounted in silicone oil. At least 300 terrestrial pollen grains and spores were identified at a minimum of 400× magnification and resolved to the lowest possible taxonomic level based on relevant atlases (Hedberg, Reference Hedberg1946; Moore et al., Reference Moore, Webb and Collinson1991; Kapp et al., Reference Kapp, Davis and King2000) and a modern reference collection at Montana State University. Coccal algae palynomorphs were enumerated from synchronous transects of pollen slides.
Pinus grains with visible, intact distal membranes were identified to subgenus. Pinus subgen. Pinus (diploxylon type, psilate distal membrane) was attributed to P. contorta-type, on the grounds that P. ponderosa does not presently grow on the Yellowstone Plateau (Arno, Reference Arno1979). Pinus subgen. Strobus (haploxylon type, verrucate distal membrane) was attributed to P. albicaulis-type, which may include P. flexilis that occurs at lower elevations in northern Yellowstone National Park. Cupressaceae grains are assigned to Juniperus-type based on the fact that only J. communis, J. horizontalis, and J. scopulorum are present in the region. Percentages of terrestrial pollen were calculated with total terrestrial pollen as a denominator, and aquatic pollen and spores were analyzed as a percent of total pollen and spores. Coccal algae were identified to the genus level (Jankovská and Komárek, Reference Jankovská and Komárek2000) and analyzed as influx ((number of individual specimens [ = NISP])/cm2/yr), given their great abundance relative to other palynomorphs. Attribution of pollen assemblages to present-day vegetation types was aided by comparison to modern pollen assemblages from the Yellowstone region (Baker, Reference Baker1976; Whitlock, Reference Whitlock1993; Iglesias et al., Reference Iglesias, Whitlock, Krause and Baker2018).
Charcoal analysis
Samples of ~2 cm3 were collected for charcoal analysis at 0.5 cm intervals for the entire composite core to reconstruct fire history. Charcoal particles >125 μm in diameter were extracted and counted according to standard methodology to analyze long-term trends in charcoal as well as local fire episodes (Whitlock and Millspaugh, Reference Whitlock and Millspaugh1996; Whitlock and Larsen, Reference Whitlock, Larsen, Smol, Birks, Last, Bradley and Alverson2001). Charcoal accumulation rates (CHAR; measured as particles/cm2/yr) were calculated with CharAnalysis software for MatLab (Higuera et al., Reference Higuera, Brubaker, Anderson, Hu and Brown2009) and were interpreted as an indicator of fire activity or overall time-averaged biomass burned.
The CHAR record was decomposed into background charcoal accumulation rates (BCHAR) representing the long-term trend, and peaks, which are positive deviations above the long-term trend (Higuera et al., Reference Higuera, Brubaker, Anderson, Hu and Brown2009, Reference Higuera, Gavin, Bartlein and Hallett2011). BCHAR was calculated using a 500-yr locally weighted scatterplot smoothing (lowess) smoother to minimize the sum of the median signal-to-noise index and the goodness-of-fit value, robust to outliers. Variations in BCHAR describe changes in regional biomass burning within tens of kilometers (Higuera et al., Reference Higuera, Whitlock and Gage2010), secondarily transported charcoal, and sediment mixing (Whitlock and Larsen, Reference Whitlock, Larsen, Smol, Birks, Last, Bradley and Alverson2001). Charcoal peaks represent fire episodes, individual fires, or a series of fires in close succession within the duration of the charcoal peak. Peaks were considered significant fire episodes if they registered above the 99th percentile of the local noise distribution of CHAR as defined by a Gaussian mixture model. Peaks were noted but considered insignificant if they registered between the 95th percentile and 99th percentile. A study of charcoal data from small lakes in the Pinus contorta forest of central Yellowstone National Park concluded that charcoal peaks register fire episodes occurring within a 1.2–3.0 km radius around the lake (Higuera et al., Reference Higuera, Whitlock and Gage2010)—a highly localized record of fire occurrence. The magnitude of a peak (particles/cm2) is generally attributed to the fire episode size (area burned) or severity. Peak detection was improved by comparison with the dates of known fires in AD 1988 and AD 1881.
Diatom analysis
Diatom samples of ~0.5 cm3 volume were collected from the core, ensuring a sample at least every 500 cal yr of the record. Sample preparation consisted of a wash with 30% wt/vol hydrogen peroxide to remove organic matter (Battarbee et al., Reference Battarbee, Jones, Flower, Cameron, Smol, Birks, Last, Bradley and Alverson2001), and mounting the residue with Naphrax optical adhesive. Diatom valves were identified to species using taxonomic resources relevant to the Northern Rocky Mountains (Bahls, Reference Bahls2005; Spaulding et al., Reference Spaulding, Bishop, Edlund, Lee, Furey, Jovanovska and Potapova2020) and general references (Patrick and Reimer, Reference Patrick and Reimer1975). A total of 300 diatom valves were enumerated per slide and analyzed as a percent of the diatom sum. Lake conditions inferred from the diatom community were based on species-level autecology.
RESULTS
Chronology
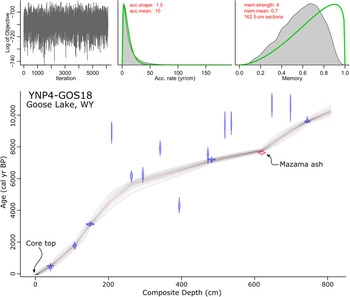
The Goose Lake age-depth model was based on 15 of 22 AMS radiocarbon ages, the age of the sediment-water interface, and the known age of Mazama ash (Egan et al., Reference Egan, Staff and Blackford2015) (Table 1, Fig. 4). Radiocarbon ages were calibrated with the IntCal13 calibration curve and verified against IntCal20 (Reimer et al., Reference Reimer, Bard, Bayliss, Beck, Blackwell, Ramsey and Buck2013, Reference Reimer, Austin, Bard, Bayliss, Blackwell, Ramsey and Butzin2020). A preliminary chronology suggested that some of the radiocarbon-dated samples were anomalously old. On the assumption that sediment accumulated over time without reversals and the age of Mazama ash was most reliable, we took a Bayesian approach to enforce a montonic model. We constrained sediment accumulation rates to positive values by setting an a priori distribution for sediment deposition time with a mean of 10 yr/cm-1 and a shape of 1.5 (Blaauw and Christen, Reference Blaauw and Christen2011). Reversals in radiocarbon ages were addressed by using Student's t-distributions with heavier tails for our calibrated ages (t a = 3, t b = 4) than for the ages of the tephra (t a = 33, t b = 44) (Christen and Pérez, Reference Christen and Pérez2009). This approach allowed us to inform the model that the ages of the known tephra were more precise than other ages and to increase their influence on the chronology accordingly (Blaauw et al., Reference Blaauw, Christen, Bennett and Reimer2018). Finally, we used splines to model depth from the explicitly combined probability distribution of all ages (Blaauw and Christen, Reference Blaauw and Christen2011).

Figure 4. Bacon age-depth model for Goose Lake composite core YNP4-GOS18. Probability distributions are plotted for the core top (light blue horizontal line), calibrated radiocarbon ages (blue distribution curves), and the Mazama ash (red distribution curve). Red dashed line is the median probability age from all run age-depth iterations, representing the best point estimate of age for any given depth, which is used for remaining plots. Gray point cloud represents age model probability and contains a 95% confidence interval (dashed gray lines). Iteration history (left top), prior and posterior densities of the mean accumulation rate (middle top), and prior and posterior of the memory (right top) suggest reasonable adherence of the model to a priori mean accumulation rate and memory assignments.
Table 1. Goose Lake core age determinations. All numbered samples were measured at NOSAMS Laboratory at the Woods Hole Oceanographic Institution. Calibrated age ranges were calculated with CALIB ver. 7.1 (Stuiver et al., 2019) and age ranges with probabilities <0.01 are excluded.

1 Calibrated age range from Egan et al. (Reference Egan, Staff and Blackford2015)
* Age outside of 95% range of final age-depth model; not used in final age model
Age reversals were present in the core below 178 cm depth, and seven radiocarbon dates were rejected by the Bacon model (Fig. 4). All but sample OS-144561 were interpreted as erroneously old. The cause of these erroneously old ages is not clear, but old carbon contamination has been documented within the Yellowstone caldera in lake sediments (Schiller et al., Reference Schiller, Whitlock, Elder, Iverson and Abbott2021) and in organic material entombed in silica sinter (Churchill et al., Reference Churchill, Manga, Hurwitz, Peek, Licciardi and Paces2020). It is likely that Goose Lake samples contained old carbon sourced from either reworked sediment, volcanic CO2 incorporated into plants during photosynthesis (Evans et al., Reference Evans, Bergfeld, McGeehin, King and Heasler2010), or volcanic CO2 in the water column and surficial sediments (Schiller et al., Reference Schiller, Whitlock, Elder, Iverson and Abbott2021).
To address the uncertainty of point estimates of age from the age-depth model, median calibrated ages with standard deviations >100 yr were rounded to the nearest 100 cal yr. The extrapolated age-depth model suggests a basal core age of 10,300 cal yr BP. A Holocene age is consistent with the fact that the basal pollen spectra were dominated by Pinus contorta-type, which was first abundant in Yellowstone Plateau pollen records after ca. 11,500 cal yr BP (Whitlock, Reference Whitlock1993; Iglesias et al., Reference Iglesias, Whitlock, Krause and Baker2018). Two changes in sediment accumulation rate were noted. Sediment accumulation rates sharply increased between 6000 cal yr BP and 4000 cal yr BP, potentially driven by a lithological change, such as the contact between Unit II and III at 178 cm depth. The change in sediment accumulation rate following Mazama ash deposition at 620 cm depth is probably an artifact of the high confidence placed on the age of the Mazama ash and the high statistical influence of the basal radiocarbon date, making age determinations below the Mazama ash potentially dubious and are aptly treated with caution hereafter.
Lithology and geochemistry
Unit I (808.5–647 cm depth; 10,300–8200 cal yr BP) consisted of light olive-brown (2.5Y 5/3), indistinctly laminated diatomaceous ooze with clay (Fig. 2). Diatoms and sponge spicules were dominant in smear slides. Diatoms were mostly intact. Peaks in sediment titanium, a non-biologically active element relatively abundant in siliciclastic sediments (Kylander et al., Reference Kylander, Ampel, Wohlfarth and Veres2011), were associated with lenses of inorganic clay, and arsenic was relatively abundant throughout the interval. δ13Csediment was measured at −18.43 ‰ VPDB.
Unit II (647–178 cm depth; 8200–3800 cal yr BP) consisted of laminated diatomaceous ooze with sapropel and minor fragmental plant remains (Fig. 2). Diatoms and sponge spicules were dominant in smear slides. Laminations were alternately grayish-brown (2.5Y 5/2), dark reddish-brown (5YR 3/2), and gray (GLEY1 5/N). Titanium peaks were associated with siliciclastic deposits, including ashes and inorganic clay layers. Arsenic was abundant throughout the unit. δ13Csediment values averaged −12.47 ‰ VPDB. An orange, inorganic clay was noted in the composite core at 628–629 cm depth (7900 cal yr BP; Fig. 3). XRD analysis indicated 81.1 wt % fluorite (hereafter fluorite mud, as an unconsolidated sediment mostly composed of inorganically precipitated fluorite), and SEM-EDS measurements confirmed abundant calcium and fluorine. The remaining weight fraction consisted of silica. SEM photomicrographs revealed diatom frustules and sponge spicules in a matrix of fluorite spherules (~0.5 μm matrix grain size, Fig. 3). The fluorite mud was notably enriched in antimony relative to the other Goose Lake sediments as indicated by XRF measurements (Fig. 3).
Two distinct siliciclastic deposits were noted in Unit II at 620–619 and 458.5–458 cm depth (Fig. 2). The lower deposit was white and composed of glass (~100%, much of which is pumiceous) with trace amounts of plagioclase, an unknown green (?)phyllosilicate, and quartz. Geochemistry of glass shards from electron microprobe analysis determined this deposit to be the Mazama ash (Iverson, N.A., personal communication, 2019). The upper deposit, which we used for correlation between cores 1A and 1D, was dark colored and a compositional mixture of lacustrine sediment, reworked minerogenic sediment, and ash. Felsic mineral abundance (siliciclastic fraction: quartz, ~60%; ash, ~40%; <1% each plagioclase, potassium feldspar, and clinopyroxene) and lacustrine components suggest a local rhyolitic source that was reworked and redeposited into Goose Lake, perhaps from surface run-off or a mass-wasting event.
Unit III (178–20.5 cm depth; 3800–200 cal yr BP) consisted of dark olive-brown (2.5Y 3/3), thinly bedded diatom ooze with sapropel (Fig. 2), containing abundant diatom frustules and chrysophytes. Biogenic sediment components, notably diatom frustules, were highly fragmented in this unit, many <2–5 μm in diameter. Highly fragmented diatom frustules suggest grazing by amphipods (e.g., Gammarus) (Quigley and Vanderploeg, Reference Quigley and Vanderploeg1991), which are abundant in modern Goose Lake (Arnold and Sharpe, Reference Arnold and Sharpe1967). δ13Csediment values were considerably lower than underlying units, −23.05 ‰ VPDB on average.
Unit IV (20.5–0 cm depth; 200 – −68 cal yr BP) consisted of very dark grayish-brown (2.5Y 3/2), massive diatomaceous sapropel with high water content (Fig. 2). Diatom frustules were relatively intact, and the δ13Csediment value was similar to the previous interval, −25.30 ‰ VPDB.
Biological proxies
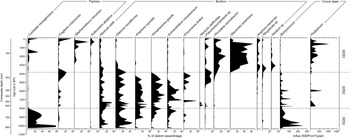
The pollen data from Goose Lake were divided into three zones based on visual inspection and using CONISS, a stratigraphically constrained cluster analysis (Grimm, Reference Grimm1987) (Fig. 5). These zones are used as a framework to describe all biological data.

Figure 5. Percentage diagrams of major pollen types and spores (>1%), total sum of terrestrial pollen, and charcoal data (CHAR and BCHAR) with significant and insignificant peaks from Goose Lake composite core YNP4-GOS18. Where present, curve exaggeration is 5×. Zone delineation is supported by CONISS dendrogram constructed with percentage data from terrestrial pollen.
Zone GOS1 (808.5–618 cm depth, 10,300–7800 cal yr BP)
Pinus dominated the pollen spectra (77%, excluding a sample immediately above the Mazama ash; Fig. 5), mostly assigned to P. contorta-type (81% of Pinus grains identifiable to subgeneric level). Artemisia (<13%), Amaranthaceae (<3%), and Poaceae (<2%) pollen were present in smaller proportions. A pollen sample immediately above the Mazama ash consisted of more Artemisia (51%) than Pinus (34%) pollen, and was the only sample from the record where the sum of nonarboreal pollen exceeded arboreal pollen types, suggesting a brief (<8 yr, based on average sediment accumulation rate for 1 cm) vegetation change following ashfall. CHAR (mean 1.4 particles/cm2/yr) was generally low (Fig. 5), although the uppermost CHAR values were probably artificially deflated by the rapid increase in sediment accumulation time noted above (Fig. 4). Mean fire frequency reached its highest values of the record (14 episodes/1000 yr).
The diatom record consisted of Cyclotella meneghiniana (28%) and Diatoma moniliformis (21%), with lesser amounts of Navicula wildii (12%), Amphora copulata (9%), Gomphonema gracile (7%), and Achnanthidium minutissimum (7%) (Fig. 6). The lower portion of Zone GOS1 was dominated solely by C. meneghiniana (Fig. 6), but by mid-zone, C. meneghiniana abundance decreased and D. moniliformis and N. wildii co-dominated. Botryococcus (89,981 NISP/cm2/yr) dominated the coccal algae assemblage with few Pediastrum (56 NISP/cm2/yr) (Fig. 6).

Figure 6. Diagram of major diatom species percentages and influx (NISP/cm2/yr) of coccal algae from Goose Lake composite core YNP4-GOS18.
Zone GOS2 (618–178 cm depth, 7750–3800 cal yr BP)
Pinus percentages increased in Zone GOS2, with abundance reaching 84% at the top of the zone (Fig. 5). P. contorta-type accounted for 93% of the identifiable Pinus. Increased Pinus percentages came at the expense of upland herb taxa, with lower abundances of Artemisia (<9%), Amaranthaceae (<2%), and Poaceae (<2%) compared to Zone GOS1. A horizon rich in degraded, indeterminate herbaceous pollen grains (457.5 cm depth, 6950 cal yr BP) was associated with the siliciclastic deposit at 458 cm depth, likely reflecting significant pollen reworking during the same depositional event. CHAR (mean 3.5 particles/cm2/yr) substantially increased from Zone GOS1 (Fig. 5), and mean fire frequency was lower than before (13 episodes/1000 yr).
The diatom record shows a loss of Cyclotella meneghiniana, and persistence of Diatoma moniliformis (17%), Gomphonema gracile (17%), Amphora copulata (16%), Navicula wildii (13%), and Achnanthidium minutissimum (12%) (Fig. 6). Additionally, an upper portion of Zone GOS2 (225–178 cm depth, 4970–3960 cal yr BP) contained a peak in Encyonema yellowstonianum (9%) and a synchronous increase in Pseudostaurosiropsis sp. (16%). The coccal algae assemblage consisted of Botryococcus (20,211 NISP/cm2/yr) and Pediastrum (3167 NISP/cm2/yr) (Fig. 6).
Zone GOS3 (178–0 cm depth, 3800– -68 cal yr BP)
Zone GOS3 was characterized by less Pinus than before (72%), predominantly P. contorta-type (95% of Pinus identifiable to subgeneric level) (Fig. 5). Major upland herb taxa concurrently increased in abundance, including Artemisia (<16%), Amaranthaceae (<3%), and Poaceae (<4%), with Ambrosia-type (<1%) and Asteraceae subfam. Asteroideae (<1%) present in low amounts. Pollen of riparian shrubs Alnus, Betula, and Salix were also present in trace amounts (<1%). Arceuthobium (<1%) was consistently detected for the first time in Zone GOS3. CHAR values (mean 9.6 particles/cm2/yr) were the lowest in the record (Fig. 5). Mean fire frequency also declined from previous zones to 10 episodes/1000 yr. The median calibrated ages from the two uppermost CHAR peaks (AD 1988–1948, AD 1903) correlate well with fires in AD 1988 and AD 1881 within LGB.
The Zone GOS3 diatom assemblage was dramatically different than previous zones and dominated by Pinnularia microstauron (14%) and Stauroneis americana (31%) (Fig. 6). Diatom abundance was lower in Zone GOS3 compared to previous zones, whereas chrysophyte cysts were more abundant. The coccal algae assemblage was dominated by Pediastrum (15,294 NISP/cm2/yr) with a lesser amount of Botryococcus (1880 NISP/cm2/yr) (Fig. 6).
DISCUSSION
In reconstructing the Holocene history of LGB, we utilize geochemical and lithological data from Goose Lake to describe changes in hydrothermal activity within and near the lake. The pollen and charcoal records are used to reconstruct the basin's vegetation and fire history, which is then compared with other sites on the Yellowstone Plateau to separate the influence of local geologic versus regional climatic drivers on ecosystem dynamics. The diatom record describes shifts in the limnobiotic conditions of Goose Lake.
Past hydrothermal activity at Goose Lake
Thermal waters flowed into Goose Lake from ca.10,300 cal yr BP to 3800 cal yr BP, based on geochemical and lithologic data. Arsenic is abundant in lithological units I and II that date to this period, and is similarly abundant in hydrothermally influenced sediments elsewhere on the Yellowstone Plateau (Shanks et al., Reference Shanks, Alt and Morgan2007; Morgan et al., Reference Morgan, Shanks and Pierce2009), as well as in present-day thermal waters in LGB (Planer-Friedrich et al., Reference Planer-Friedrich, Lehr, Matschullat, Merkel, Nordstrom and Sandstrom2006). In addition, antimony-rich fluorite mud, deposited briefly ca. 7900 cal yr BP (619–618 cm depth), is definitively of hydrothermal provenance, inasmuch as fluorite is a secondary hydrothermal mineral in the rhyolite flows that underlie Goose Lake (Bargar and Beeson, Reference Bargar and Beeson1981), and antimony is abundant in geothermal waters from LGB (Stauffer and Thompson, Reference Stauffer and Thompson1984). Lithologic units I and II also have higher δ13Csediment values (−18.43 ‰ VPDB; −9.7 ‰ VPDB) than overlying units, pointing to an admixture of hydrothermally degassed carbon (~−3 ‰ VPDB; Bergfeld et al., Reference Bergfeld, Lowenstern, Hunt, Shanks and Evans2014), which is higher than terrestrial vegetation (Smith and Epstein, Reference Smith and Epstein1971), and terrestrial or aquatic organic carbon. In addition, the more distinct laminations (Fig. 2) of sediments in Unit I and, especially, Unit II suggest frequently shifting limnological conditions as might be produced by intermittent inputs of thermal water relative to meteoric and groundwater sources. Sediments remain lacustrine in character with abundant diatom frustules and sponge spicules in Units I and II. Thus, Goose Lake during this interval was adjacent to or supported sublacustrine thermal features. The presence of a benthic lacustrine community indicates that it was not a large thermal spring, such as present-day Firehole Lake or Hot Lake in LGB (Fig. 1B).
Goose Lake hydrothermal activity ceased abruptly at ca. 3800 cal yr BP. Diminished arsenic abundance, lower 13Csediment values (−25.30 ‰ VPDB; −22.08 ‰ VPDB), and darker sediments composed of diatoms and chrysophytes in lithologic Units III and IV suggest quiescent lake sedimentation without hydrothermal influence (Fig. 2). The abruptness of this transition is inferred from the sharp contact between lithologic Units II and III.
Development of the LGB geo-ecosystem with regional context
To determine the relative importance of local hydrothermal activity versus regional climate change in shaping the environmental history of LGB, we compared the Goose Lake data with three other paleoecological records from the Yellowstone Plateau. We assumed that concurrent and similar paleoecological changes across many sites point to a regional forcing, such as climate change. When a vegetation change occurred only at one site, the forcing was likely local, such as changing hydrothermal activity.
The vegetation record at Goose Lake was compared with nearby sites that lack significant surficial hydrothermal activity: Cygnet Lake (Whitlock, Reference Whitlock1993; Millspaugh et al., Reference Millspaugh, Whitlock and Bartlein2000), Loon Lake (Whitlock et al., Reference Whitlock, Bartlein and Van Norman1995), and Yellowstone Lake (Theriot et al., Reference Theriot, Fritz, Whitlock and Conley2006) (Fig. 1A, Table 2). Yellowstone Plateau forests are dominated by Pinus contorta, but the admixture of Pseudotsuga menziesii at Loon Lake and Abies lasiocarpa and Picea engelmannii at Yellowstone Lake necessitates acknowledgement of other arboreal forest components. Therefore, we used a standardized ratio of arboreal pollen (AP) to non-arboreal pollen (NAP) types (AP:NAP, AP – NAP/AP + NAP) to infer past forest cover (openness or extent) at each site. A lowess smoother (x ~ y) was applied to each AP:NAP data set to emphasize long-term trends.
Table 2. Paleoecological sites and reference sources from within Pinus contorta forest site from Yellowstone National Park and the Yellowstone Plateau (an EPA Level IV Ecoregion; Chapman et al., Reference Chapman, Bryce, Omernik, Despain, ZumBerge and Conrad2004) used for comparison purposes with Goose Lake data.

*Source of pollen data
† Source of charcoal data
A charcoal-based fire history at Cygnet Lake provides a reasonable comparison for that at Goose Lake, given that the site is similar in size and also surrounded by Pinus contorta forest (Millspaugh et al., Reference Millspaugh, Whitlock and Bartlein2000). CharAnalysis was conducted for Cygnet Lake charcoal data in the same manner as for Goose Lake, except BCHAR values at Cygnet Lake were calculated with the use of a 700-yr lowess smoother robust to outliers, which minimized the sum of the median signal-to-noise index and the goodness-of-fit value. New age models were created for Cygnet, Loon, and Yellowstone Lakes utilizing IntCal13 (Reimer et al., Reference Reimer, Bard, Bayliss, Beck, Blackwell, Ramsey and Buck2013) and Bacon (Blaauw and Christen, Reference Blaauw and Christen2011), and sediment accumulation rate priors that closely approximated inferences from the original publications (Whitlock, Reference Whitlock1993; Whitlock et al., Reference Whitlock, Bartlein and Van Norman1995; Millspaugh et al., Reference Millspaugh, Whitlock and Bartlein2000; Theriot et al., Reference Theriot, Fritz, Whitlock and Conley2006).
Holocene climate context on the Yellowstone Plateau
The Holocene climate of the Yellowstone Plateau was strongly influenced by slow variations in the seasonal cycle of precession, which led to a summer insolation maximum in the Northern Hemisphere at ca. 11 ka and a steady decline thereafter (Laskar et al., Reference Laskar, Robutel, Joutel, Gastineau, Correia and Levrard2004). The Yellowstone Plateau has a summer-dry precipitation regime (Whitlock and Bartlein, Reference Whitlock and Bartlein1993), and the summer insolation maximum (summer anomaly up to ~30 W/m2 at 44.5°N; Laskar et al., Reference Laskar, Robutel, Joutel, Gastineau, Correia and Levrard2004) in this region resulted in warmer and effectively drier conditions than the twentieth century average as a result of the expansion and strengthening of the northeastern Pacific subtropical high-pressure system (Whitlock and Bartlein, Reference Whitlock and Bartlein1993; Bartlein et al., Reference Bartlein, Anderson, Anderson, Edwards, Mock, Thompson, Webb, Iii and Whitlock1998). In the middle and late Holocene, summers on the Yellowstone Plateau became cooler and effectively wetter due to declining summer insolation and the attendant weakening of the northeastern Pacific subtropical high, leading to the establishment of present climate conditions (Whitlock and Bartlein, Reference Whitlock and Bartlein1993; Bartlein et al., Reference Bartlein, Anderson, Anderson, Edwards, Mock, Thompson, Webb, Iii and Whitlock1998). Paleoclimate reconstructions from the Yellowstone National Park vicinity corroborate a story of warm, dry conditions in the early Holocene that became increasingly cool and wet to the present (Larsen et al., Reference Larsen, Crump and Blumm2020; Chellman et al., Reference Chellman, Pederson, Lee, McWethy, Puseman, Stone, Brown and McConnell2021).
Early Holocene (GOS1, >10,300–7800 cal yr BP)
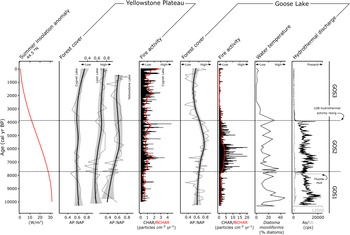
Pollen records from Cygnet Lake, Loon Lake, and Yellowstone Lake indicate that surrounding forests were relatively open or less extensive, as evidenced by depressed AP:NAP values at all sites compared with the Holocene average from the area (Fig. 7; Whitlock, Reference Whitlock1993; Whitlock et al., Reference Whitlock, Bartlein and Van Norman1995; Theriot et al., Reference Theriot, Fritz, Whitlock and Conley2006). The landscape at Goose Lake was similarly open, as indicated by similarly low AP:NAP (Fig. 7). Forests around Goose Lake were dominated by Pinus contorta as they are at present, as indicated by a dominance of Pinus pollen (Fig. 5). The deposition of the Mazama ash at 7630 cal yr BP (Egan et al., Reference Egan, Staff and Blackford2015) led to a short-term proliferation of Artemisia, as evidenced by the spike in Artemisia percentages immediately above the ash. This spike is attributed to short-lived mulching effects of volcanic ash on the landscape, and has been noted in records throughout the northern Rocky Mountain region (Schiller et al., Reference Schiller, Whitlock, Alt and Morgan2020). The charcoal record from Cygnet Lake shows that fire activity was generally low in the early Holocene as evidenced by the lowest CHAR of the record (mean CHAR 0.53 particles/cm2/yr). Fire activity at Goose Lake was similarly low in this period compared with the rest of the record based on BCHAR data, and fire frequency was the highest of the record (Fig. 7). Fires were frequent but relatively small or low in severity.

Figure 7. Comparison plot of proxy data from the Goose Lake composite core YNP4-GOS18 with other sites on the Yellowstone Plateau (Cygnet Lake, Loon Lake, Yellowstone Lake). Seasonal insolation anomaly calculated for 44.5°N (Laskar et al., Reference Laskar, Robutel, Joutel, Gastineau, Correia and Levrard2004). Forest cover at sites without significant surficial hydrothermal activity (Whitlock, Reference Whitlock1993; Whitlock et al., Reference Whitlock, Bartlein and Van Norman1995; Theriot et al., Reference Theriot, Fritz, Whitlock and Conley2006) is plotted as standardized AP:NAP ratios (AP - NAP/AP + NAP; dotted line) with lowess smoother (solid line with shaded 95% confidence interval); fire activity at Cygnet Lake (Millspaugh et al., Reference Millspaugh, Whitlock and Bartlein2000; CHAR in black, BCHAR in red). Forest cover and fire activity plots for Goose Lake are juxtaposed with diatom and geochemical indicators of hydrothermal conditions.
The diatom data indicate that Goose Lake was shallower than at present, with oligotrophic to mesotrophic conditions in the early Holocene. Cyclotella meneghiniana, Amphora copulata, and Achnanthidium minutissimum are favored by oligotrophic to mesotrophic conditions at present (Parsons et al., Reference Parsons, Dortch, Turner and Rabalais2006; Salm et al., Reference Salm, Saros, Martin and Erickson2009; Saros and Anderson, Reference Saros and Anderson2015). C. meneghiniana populations are abundant in well-mixed or relatively shallow lakes (Reynolds, Reference Reynolds1980), and bloom in response to influxes of nitrogen and phosphorous (Salm et al., Reference Salm, Saros, Martin and Erickson2009). The presence of Diatoma moniliformis is consistent with water temperatures greater than 10–15°C (Pniewski and Sylwestrzak, Reference Pniewski and Sylwestrzak2018) and/or harsh conditions (e.g., the species is found in nuclear power-plant cooling channels; Potapova and Snoeijs, Reference Potapova and Snoeijs1997). Another benthic species, Navicula wildii, prefers neutral to moderate alkalinity (Bahls, Reference Bahls2012), and Achnanthidium minutissimum is present in a wide variety of pH conditions (Ponader and Potapova, Reference Ponader and Potapova2007). Gomphonema gracile is a cosmopolitan benthic species (Patrick and Reimer, Reference Patrick and Reimer1975). The assemblage, along with its good preservation (i.e., diatoms are destroyed under strongly alkaline conditions) indicates that the water pH was neutral to slightly alkaline, similar to that of most LGB thermal waters at present (Hurwitz and Lowenstern, Reference Hurwitz and Lowenstern2014). The dominance of C. meneghiniana from 10,300 cal yr BP to 9800 cal yr BP suggests an initial period of high nutrient input, and the later abundance of low-nutrient specialist Amphora copulata implies decreasing nutrient availability after 9420 cal yr BP.
Botryococcus was abundant in Goose Lake in the early Holocene and Pediastrum was nearly absent; an assemblage often interpreted as evidence of extremely cold, oligotrophic, or acidic waters (Jankovská and Komárek, Reference Jankovská and Komárek2000). Those conditions seem unlikely at this time in Goose Lake in light of the diatom assemblage, which suggests neutral to alkaline water chemistry and inferred warm, dry summers (Whitlock and Bartlein, Reference Whitlock and Bartlein1993). However, high levels of arsenic in the water column may explain the Botryococcus abundance. Some modern waterways contaminated by anthropogenic (Meeinkuirt et al., Reference Meeinkuirt, Sirinawin, Angsupanich and Polpunthin2008) or naturally occurring (Miller, Reference Miller2020) arsenic have abundant Botryococcus, with little to no noted Pediastrum. B. braunii has even shown some promise as a bioremediator of environmental arsenic (Onalo et al., Reference Onalo, Matias-Peralta and Sunar2014; Podder and Majumder, Reference Podder and Majumder2016). Taken together, the diatom and coccal algae assemblages indicate warm, somewhat alkaline, and arsenic-rich conditions associated with thermal water discharge into Goose Lake.
In summary, early Holocene vegetation at Goose Lake resembled that of the Yellowstone Plateau as a whole, with relatively open Pinus contorta forest or expanded non-forested vegetation as a result of warm, dry conditions and frequent small or low-severity fires. Thermal waters discharged directly into Goose Lake, as evidenced by the geochemical and lithological characteristics of the sediments and limnobiotic communities, but hydrothermal activity did not alter the upland vegetation to a large extent.
Middle Holocene (GOS2, 7750–3800 cal yr BP)
Increasing AP:NAP values from Cygnet Lake, Loon Lake, and Yellowstone Lake suggest that forest became more closed or more extensive across the Yellowstone Plateau in the middle Holocene (Whitlock, Reference Whitlock1993; Whitlock et al., Reference Whitlock, Bartlein and Van Norman1995; Theriot et al., Reference Theriot, Fritz, Whitlock and Conley2006; Fig. 7). At Goose Lake, the forest also became more closed or extensive through this period, with similarly increasing AP:NAP values (Fig. 7) and continued dominance of Pinus pollen (Fig. 5). An increase in fire activity at Cygnet Lake, as evidenced by rising BCHAR, was coupled with particularly extreme (large or severe) fire episodes based on high charcoal peak magnitudes (Millspaugh et al., Reference Millspaugh, Whitlock and Bartlein2000). The same interpretation can be drawn at Goose Lake, where the charcoal data indicate large or severe, but infrequent, fires (Fig. 7).
The middle Holocene diatom assemblage at Goose Lake is similar to that of the early Holocene, except for the conspicuous absence of Cyclotella meneghiniana. The dominance of benthic taxa suggests a change in water level, but it is difficult to know if water level rose or fell, given the broad, flat bathymetry of Goose Lake (Fig. 1C). Persistence of species with warm-water preferences (Diatoma moniliformis) and wide pH tolerance ranges (Achnanthidiuim minutissimum, Navicula wildii, and Pseudostaurosiropsis sp.) (Ciniglia et al., Reference Ciniglia, Cennamo, De Stefano, Pinto, Caputo and Pollio2007) indicates that the diatom assemblage continued to be influenced by thermal inputs through this period. Low nutrient availability is indicated by Achnanthidiuim minutissimum and Amphora copulata. Peaks of Encyonema yellowstonianum and Pseudostaurosiropsis sp., both colonial tube-forming taxa, suggest periods of enhanced turbidity (Krammer, Reference Krammer1997; Ciniglia et al., Reference Ciniglia, Cennamo, De Stefano, Pinto, Caputo and Pollio2007). Botryococcus and Pediastrum both occur in this interval, suggesting improved limnic conditions for Pediastrum growth.
In summary, during the increasingly cool, wetter summers of the middle Holocene, Pinus contorta forest at Goose Lake continued to resemble that of other sites on the Yellowstone Plateau, with increasingly closed or extensive Pinus contorta forest and greater fire activity. Geochemical and limnobiotic proxies indicate a sustained thermal influence at Goose Lake, and the diatom data suggest shifts in lake level and enhanced turbidity.
Late Holocene, modern Lower Geyser Basin (GOS3, 3800 cal yr BP–present)
AP:NAP values from Cygnet Lake and Loon Lake suggest continued forest expansion on the Yellowstone Plateau (Whitlock, Reference Whitlock1993; Whitlock et al., Reference Whitlock, Bartlein and Van Norman1995). At Yellowstone Lake, AP:NAP values remain stable, suggesting little change in forest cover at a regional scale (Theriot et al., Reference Theriot, Fritz, Whitlock and Conley2006). In contrast, the forest at Goose Lake became more open or restricted, as evidenced by decreased AP:NAP values (Fig. 7) resulting from an increase in Artemisia, Poaceae, and Amaranthaceae percentages (Fig. 5). This shift likely marks the establishment of modern, open vegetation in LGB. Artemisia and Amaranthaceae are not abundant in LGB today and the pollen may have come from steppe vegetation in other parts of Yellowstone National Park, or the Snake River Plain, Idaho, which lies upwind to the west. Both Artemisia and Amaranthaceae are prolific pollen producers and long-distance transport from low elevation, semiarid regions has been well documented in pollen studies from the region (Fall, Reference Fall1992; Whitlock, Reference Whitlock1993; Iglesias et al., Reference Iglesias, Whitlock, Krause and Baker2018). Steady but low amounts of Alnus, Betula, and Salix pollen in the late Holocene suggest the presence of riparian areas and wet meadows near Goose Lake (Fig. 5). Charcoal levels at Cygnet Lake indicate increasing fire activity and infrequent, large fire episodes (Fig. 7). In contrast, fire activity declined at Goose Lake, suggesting a more open landscape with little fuel biomass. Fire episodes, evidenced by charcoal peaks, were also less frequent than before at Goose Lake (Fig. 5).
The late Holocene diatom assemblage at Goose Lake differs from that of previous periods, and suggests that the water column was well-mixed with relatively low conductivity. The dominant taxa were Stephanodiscus minutulus, Pinnularia microstauron, and Stauroneis americana. Stephanodiscus minutulus is a planktic, high-phosphorous specialist with low nitrogen and silica requirements that blooms in early spring during isothermal mixing (Lynn et al., Reference Lynn, Kilham, Kreeger and Interlandi2000; Interlandi et al., Reference Interlandi, Kilham and Theriot2003; Theriot et al., Reference Theriot, Fritz, Whitlock and Conley2006). P. microstauron is a benthic species that favors oligotrophic, oxygen-rich waters with low conductivity (Krammer, Reference Krammer and Lange-Bertalot2000). Stauroneis americana is benthic, preferring wetlands with low to moderate conductivity (Vijver et al., Reference Vijver, Gremmen and Beyens2005). Pediastrum dominated the coccal algae assemblage at Goose Lake in this interval, probably growing along the shallow margins of the lake despite oligotrophic to mesotrophic conditions.
In summary, we suggest an abrupt reorganization of hydrothermal activity in LGB at ca. 3800 cal yr BP explains the pollen and diatom records at Goose Lake (Fig. 7). The Goose Lake pollen record diverges from other Yellowstone Plateau records by showing evidence of more open or limited forest during the late Holocene at a time when forests were becoming more closed or extensive elsewhere in the region. The presence of diatom taxa that previously thrived in warm, thermally influenced waters declined, and the Botryococcus-dominated coccal algae assemblage was almost fully replaced by Pediastrum. We hypothesize that the development of open vegetation and changes in limnobiota mark the creation of the modern, broad, thermal flat in LGB. This change was contemporaneous with the cessation of hydrothermal activity at Goose Lake.
Potential mechanisms of geo-ecological change ca. 3800 cal yr BP
The shift in hydrothermal activity away from the lake and synchronous creation of broad thermal flats at ca. 3800 yr BP seem a likely explanation for the abiotic and biotic changes registered at Goose Lake, but there is no independent evidence to corroborate this reconstruction. LGB thermal features have not been directly dated, and organic material entombed in siliceous sinter deposits in Upper Geyser Basin ~6 km south of LGB either do not date beyond the past thousand years (Marler, Reference Marler1956; Hurwitz et al., Reference Hurwitz, King, Pederson, Martin, Damby, Manga, Hungerford and Peek2020), or are unreliable (Lowenstern et al., Reference Lowenstern, Hurwitz and McGeehin2016; but see Churchill et al., Reference Churchill, Manga, Hurwitz, Peek, Licciardi and Paces2020).
The shift in hydrothermal activity at 3800 yr BP may have been triggered by changes in geology or climate. Earthquakes (Miller, Reference Miller1968; Marler and White, Reference Marler and White1975; Pitt and Hutchinson, Reference Pitt and Hutchinson1982) and ground deformation (Vaughan et al., Reference Vaughan, Hungerford and Keller2020) are known to abruptly alter hydrothermal activity, and these events have probably affected LGB hydrothermal activity throughout the Holocene. For comparison, the 1959 Hebgen Lake Earthquake (M 7.1, epicenter ~40 km from the Yellowstone Plateau's western thermal areas) altered the activity of many existing thermal features (Marler and White, Reference Marler and White1975) and caused new soil heating in the vicinity of Rush Lake in LGB, destroying adjacent forest (Miller, Reference Miller1968). An earthquake at ca. 3800 cal yr BP could have caused the extinction of the thermal feature(s) at Goose Lake. A significant fault along the western shore of Yellowstone Lake is dated to ca. 3.3 ka based on scarp morphology, radiocarbon dating, and obsidian hydration dating (Locke et al., Reference Locke, Meyer and Pings1992). Ground uplift related to inflation of the Yellowstone caldera also occurred after ca. 3.8 ka, as inferred from drowned streams and radiocarbon dates at Yellowstone Lake (Pierce et al., Reference Pierce, Cannon, Meyer, Trebesch, Watts and Morgan2007a). Either a single or combination of such geological forcings may have been responsible for the hydrothermal shift noted in the Goose Lake record.
Alternatively, climate-driven changes in hydrology may have altered LGB hydrothermal activity at 3800 cal yr BP. For example, Old Faithful Geyser ceased to erupt during a multidecadal drought in the Medieval Climate Anomaly (Hurwitz et al., Reference Hurwitz, King, Pederson, Martin, Damby, Manga, Hungerford and Peek2020), so a loss of effective moisture might have altered LGB hydrothermal activity similarly. However, the gradual increase in effective moisture in the late Holocene (Larsen et al., Reference Larsen, Crump and Blumm2020; Chellman et al., Reference Chellman, Pederson, Lee, McWethy, Puseman, Stone, Brown and McConnell2021) seems inconsistent with abrupt reorganization of the LGB hydrothermal system. Reduced precipitation, however, would have altered the subsurface water supply of LGB, and may have resulted in a reorganization of its hydrothermal activity, including cessation of hydrothermal activity at Goose Lake. This change may have been caused by abrupt drought conditions correlated with the 4.2 ka climate event, a multidecadal period of severe drought in North America (Booth et al., Reference Booth, Jackson, Forman, Kutzbach, Bettis, Kreigs and Wright2005; Lundeen et al., Reference Lundeen, Brunelle, Burns, Polyak and Asmerom2013; Carter et al., Reference Carter, Shinker and Preece2018) and much of the Northern Hemisphere (Railsback et al., Reference Railsback, Liang, Brook, Voarintsoa, Sletten, Marais, Hardt, Cheng and Edwards2018). An ice core described from the Beartooth Mountains northeast of Yellowstone National Park indicated a period of anomalous, warmer winter-season temperatures between 4200 cal yr BP and 3800 cal yr BP (Chellman et al., Reference Chellman, Pederson, Lee, McWethy, Puseman, Stone, Brown and McConnell2021), which may have led to drier conditions in the region. In any case, there has been difficulty in attributing the cause of changes to modern hydrothermal features (e.g., Reed et al., Reference Reed, Munoz-Saez, Hajimirza, Wu, Barth, Girona and Rasht-Behesht2021), which cautions against hasty, causal attribution of such changes, particularly in the deep past.
CONCLUSIONS
The Yellowstone Plateau and surrounding mountainous regions have been termed a geo-ecosystem because the ecosystem and other environmental factors are strongly influenced by geological processes, including volcanism, hydrothermal activity, faulting, and tectonic uplift (Pierce et al., Reference Pierce, Despain, Morgan and Good2007b). Paleoecological studies in other regions have noted the legacy of geological processes in shaping vegetation history, either in placing edaphic constraints on vegetation development (Brubaker, Reference Brubaker1975; Bernabo, Reference Bernabo1981; Briles et al., Reference Briles, Whitlock, Skinner and Mohr2011) or in triggering sudden ecological disturbances (Ogden et al., Reference Ogden, Fordham, Horrocks, Pilkington and Serra2005; Boyd et al., Reference Boyd, Lentfer and Parr2005; Long et al., Reference Long, Power, Minckley and Hass2014). In most situations, the driving geologic event(s) usually occurred in the deep past (e.g., formation and exhumation of infertile, ultramafic substrate) or had a brief ecological impact (e.g., most volcanic ashfalls). By contrast, the Yellowstone geo-ecosystem remains active, and its ecological history shows a dynamic response to past and ongoing geological processes in concert with climate change. The Holocene record at Goose Lake provides a glimpse into the nature of these interactions at Lower Geyser Basin, with a marked shift in hydrothermal activity during the last ca. 10,300 cal yr that affected the terrestrial and aquatic ecosystems.
Thermal waters discharged directly into Goose Lake prior to ca. 3800 cal yr BP and had little influence on upland vegetation in the basin. In early and middle Holocene, the Pinus contorta forest of LGB became increasingly closed and fire activity was generally high, as observed at other sites on the Yellowstone Plateau. Limnic conditions, however, were affected by thermal water discharge, as evidenced by the presence of diatom and coccal algae assemblages adapted to extreme, warm, and potentially somewhat alkaline conditions.
An abrupt reorganization of hydrothermal activity is inferred ca. 3800 cal yr BP, when thermal waters no longer fed directly into Goose Lake. A seismic event may have triggered this shift in basin hydrothermal activity, just as recent earthquakes and uplift have done.
Hydrothermal activity may have become more widespread in LGB ca. 3800 cal yr BP, as inferred from expansion of non-forest vegetation types. The late Holocene vegetation and fire history at Goose Lake diverges from other sites in the region by indicating that the area became more open and sustained few fires. This configuration marks the development of the modern, treeless thermal flat.
Acknowledgments
Field work was conducted under Yellowstone National Park permit YELL-2018-SCI-0009. A. Carlson, R.E. Gresswell, N.E. Kichas, and W.P. Nanavati helped with field work, and J.A. Eggers, B. Hosseini, D.W. Mogk, N. Rieders, and E. Roehm provided laboratory assistance. N.A. Iverson provided unpublished tephrochronological data, V. Iglesias helped with the age-depth model, and S.C. Fritz reviewed the diatom interpretations. S. Hurwitz, J.B. Lowenstern, L.A. Morgan, and an anonymous reviewer provided helpful reviews of the manuscript.
Financial Support
This research was funded by NSF Grant EAR-1515353 to C. Whitlock as part of the HD-YLAKE project and a research fellowship from the Montana Institute on Ecosystems to C.M. Schiller. This work was performed in part at the Montana Nanotechnology Facility, a member of the National Nanotechnology Coordinated Infrastructure (NNCI), which is supported by the NSF Grant ECCS-1542210.





 Open access
Open access