



Introduction
The classic palaeontological locality of Nahoryany (Nagórzany in Polish literature), south of Lviv in western Ukraine (Fig. 1), has long been known as a rich source of spectacular macrofossils of early Maastrichtian marine invertebrates. In the nineteenth century, these were sent out to many European museums. The first account of this locality and its fossils was published in the newspaper ‘Gazeta Lwowska’ (Kronenthal, 1843). The reader is referred to Kennedy & Summesberger (Reference Kennedy and Summesberger1987) and Machalski & Malchyk (Reference Machalski and Malchyk2016) for historical overviews of the Nahoryany section. Today, it is impossible to collect fossils at Nahoryany, because the local quarries were abandoned near the end of the nineteenth century (Łomnicki, Reference Łomnicki1897). However, rich materials from this site are available for study in museum collections both in Ukraine and in western Europe (e.g., Kotsiubynskyi, Reference Kotsiubynskyi1958, Reference Kotsiubynskyi, Pasternak, Havrylishyn, Hynda, Kotsiubynskyi and Senkovskyi1968; Christensen, Reference Christensen1987; Kennedy & Summesberger, Reference Kennedy and Summesberger1987; Bakayeva, Reference Bakayeva2011; Machalski & Malchyk, Reference Machalski and Malchyk2016).

Figure 1. Location of the Nahoryany site in western Ukraine.
Scaphitid ammonites, that is, representatives of the family Scaphitidae Gill, Reference Gill1871, are a characteristic component of the Nahoryany fauna, having been studied and/or discussed in several papers (e.g., Kner, Reference Kner1848, Reference Kner1852; Alth, Reference Alth1850; Geinitz, 1850; Favre, Reference Favre1869; Mikhailov, Reference Mikhailov1951; Błaszkiewicz, Reference Błaszkiewicz1980; Kennedy & Summesberger, Reference Kennedy and Summesberger1987; Kennedy, Reference Kennedy and House1993; Machalski & Odin, Reference Machalski, Odin and Odin2001; Machalski, Reference Machalski and Bogdanova2019). Representatives of the two lineages distinguished by Machalski (Reference Machalski and Bogdanova2019) in the Maastrichtian of Europe are present in the Nahoryany collections. The first lineage comprises giant scaphitids, leading from Hoploscaphites quadrispinosus (Geinitz, 1850) through Acanthoscaphites (Acanthoscaphites) tridens (Kner, Reference Kner1848) to A. (Euroscaphites) varians blaszkiewiczi Jagt, et al., Reference Jagt, Kennedy and Machalski1999, and A. (E.) v. varians (Łopuski, Reference Łopuski1911). Both H. quadrispinosus and A. (A.) tridens occur at Nahoryany (Machalski & Malchyk, Reference Machalski and Malchyk2016). Another lineage, the focus of the present paper, comprises Hoploscaphites pumilus (Stephenson, Reference Stephenson1941), which led to H. constrictus (J. Sowerby, Reference Sowerby1817); this is here referred to as the Hoploscaphites constrictus lineage. The eponymous species is a common European taxon, established on early late Maastrichtian material from the Calcaire à Baculites of the Cotentin Peninsula, Manche, north-west France (see Kennedy, Reference Kennedy1986; Walaszczyk & Kennedy, Reference Walaszczyk and Kennedy2011). A group of mid- to late Maastrichtian taxa, e.g., Hoploscaphites tenuistriatus (Kner, Reference Kner1848) or H. schmidi (Birkelund, Reference Birkelund1982), are interpreted as short-lived offshoots of the main H. constrictus lineage (Machalski, Reference Machalski2005). Additionally, there are also rare records of North American immigrants from the European Maastrichtian (Jagt & Kennedy, Reference Jagt and Kennedy1994; Machalski et al., Reference Machalski, Jagt, Landman and Motchurova-Dekova2007).
In the past, Hoploscaphites constrictus was regarded as an informal index fossil for the Boreal Maastrichtian in Europe, having been even considered as one of the potential markers for the lower boundary of the stage as understood then (e.g., Schmid, Reference Schmid1955; Birkelund et al., Reference Birkelund, Hancock, Hart, Rawson, Remane, Robaszynski, Schmid and Surlyk1984; Kennedy, Reference Kennedy1984b; Schulz et al., Reference Schulz, Ernst, Ernst and Schmid1984). Reference is made to Walaszczyk et al., (Reference Walaszczyk, Cobban and Odin2002a, Reference Walaszczyk, Odin and Dhondt2002b), Niebuhr et al. (Reference Niebuhr, Hampton, Gallagher and Remin2011), Machalski (Reference Machalski2012a), Remin (Reference Remin2012), Walaszczyk et al. (Reference Walaszczyk, Dubicka, Olszewska-Nejbert and Remin2016) and Wilmsen et al. (Reference Wilmsen, Engelke, Linnert, Mutterlose and Niebuhr2018) for correlations between the traditional position for the base of the Maastrichtian, which coincided with the base of the conventional belemnite Belemnella lanceolata Zone (e.g., Christensen, Reference Christensen1996), with its currently accepted location. The latter has been based on the Global Stratotype Section and Point for the lower boundary of Maastrichtian Stage at a level 115.2 m in the depositional succession exposed at Tercis les Bains, Landes, south-west France (Odin & Lamaurelle, Reference Odin and Lamaurelle2001).
Hoploscaphites constrictus has also been claimed to represent an example of evolutionary stasis (Kennedy, Reference Kennedy1989). However, subsequent studies have revealed that records from the lowermost Maastrichtian are, in fact, either based on H. pumilus or doubtful (Machalski & Odin, Reference Machalski, Odin and Odin2001; Machalski, Reference Machalski and Bogdanova2019). It has also been demonstrated that younger representatives of the H. constrictus lineage may be subdivided into temporal subspecies, with the last one, H. constrictus johnjagti Machalski, Reference Machalski2005, briefly surviving Cretaceous–Paleogene boundary event (Machalski, Reference Machalski2005; Machalski & Heinberg, Reference Machalski and Heinberg2005; Machalski, Reference Machalski, Jagt and Jagt-Yazykova2012b).
Machalski & Odin (Reference Machalski, Odin and Odin2001) proposed that Hoploscaphites constrictus descended, via transitional H. constrictus anterior Błaszkiewicz, Reference Błaszkiewicz1980, from H. pumilus, a species described originally from the upper Campanian of North America (Stephenson, Reference Stephenson1941; Cobban, Reference Cobban1974; Kennedy & Cobban, Reference Kennedy and Cobban1993), but subsequently recorded also from the upper Campanian and lower Maastrichtian in France (Kennedy et al., Reference Kennedy, Bilotte, Lepicard and Segura1986; Machalski & Odin, Reference Machalski, Odin and Odin2001; Odin et al., Reference Odin, Courville, Machalski, Cobban and Odin2001). Discussions on that subject also involved specimens from Nahoryany, but results were not satisfactory. This is exemplified by a suite of specimens from Nahoryany assigned to Scaphites constrictus by Favre (Reference Favre1869, pl. 5, figs. 1, 2, 4), to Hoploscaphites constrictus by Kennedy & Summesberger (Reference Kennedy and Summesberger1987), tentatively affiliated with H. c. anterior by Błaszkiewicz (Reference Błaszkiewicz1980), and with H. pumilus and H. anterior (raised to species level) by Machalski & Odin (Reference Machalski, Odin and Odin2001).
In view of all the above, the aim of the present paper is twofold: (1) to reassess the taxonomy of specimens of the genus Hoploscaphites Nowak, Reference Nowak1911 from Nahoryany and (2) to discuss the evolutionary origin and stratigraphical range of Hoploscaphites constrictus.
Geological setting
There is limited data on the exact location and appearance of the Nahoryany outcrop, of which there are no traces in the present landscape as checked by myself in 2001. According to Kronenthal (1843), the Nahoryany section was located in a forest, two miles south of Lviv, on the right side of the road leading to the town of Stryj. The section was available in two quarries situated close to each other. At the quarries marls, limestones and sandstones were reportedly exposed; these yielded abundant fossils of marine invertebrates, including often large-sized cephalopod and bivalve specimens, which occasionally formed accumulations. Kner (Reference Kner1848), mentioned two quarries of a few ‘Klafter’ depths exposing alternating sandstones and limestones; the Viennese Klafter was equal to 1.896 m.
During examination of Nahoryany specimens, I did not come across any sandstone matrix; the ‘sandstones’ probably represented sandy limestones and marls. In Polish literature, the Nahoryany rocks were commonly referred to as the ‘opoka nagórzańska’ (‘Nahoryany opoka’, see e.g., Łomnicki, Reference Łomnicki1897; Siemiradzki, Reference Siemiradzki1905; Syniewska, Reference Syniewska1923). Opoka is a carbonate-siliceous rock that is widely distributed in Upper Cretaceous successions across Poland and Ukraine, but according to Jurkowska & Świerczewska-Gładysz (Reference Jurkowska and Świerczewska-Gładysz2022), its proper identification requires a petrological and mineralogical study.
The environmental setting of the Nahoryany succession has never been the subject of analysis. However, the admixture of sand, the abundance of large-sized molluscs and the presence of fossil accumulations point to deposition in a relatively shallow-water, turbulent environment (see Machalski & Malchyk, Reference Machalski and Malchyk2019, figure 3, for a facies model of Upper Cretaceous deposits in Poland and western Ukraine).
As far as the stratigraphy of the section is concerned, there are overviews in Mikhailov (Reference Mikhailov1951), Pasternak et al., (Reference Pasternak, Havrylyshyn, Hynda, Kotsiubynskyi and Senkovskyi1968, Reference Pasternak, Senkovskyi and Havrylyshyn1987), Kennedy & Summesberger (Reference Kennedy and Summesberger1987) and Machalski & Malchyk (Reference Machalski and Malchyk2016). According to Pasternak et al., (Reference Pasternak, Havrylyshyn, Hynda, Kotsiubynskyi and Senkovskyi1968, Reference Pasternak, Senkovskyi and Havrylyshyn1987), the Nahoryany section belonged to the ‘Poteleckaja svita’ of the lower Maastrichtian, characterised by the occurrence of ‘Belemnella lanceolata and Acanthoscaphites tridens’. These deposits are overlain by the chalk succession of the ‘Lvivskaja svita’ (‘opoka lwowska’ or ‘Lviv opoka’ of authors) assigned to the conventional lower upper Maastrichtian Belemnitella junior Zone. See Christensen (Reference Christensen1996, fig. 1) for conventional belemnite zonation of the Maastrichtian and Machalski & Malchyk (Reference Machalski and Malchyk2016, fig. 4) for belemnite and inoceramid zonation of the upper Campanian and Maastrichtian successions in western Ukraine.
Inoceramid data published by Kotsiubynskyi (Reference Kotsiubynskyi1958, Reference Kotsiubynskyi, Pasternak, Havrylishyn, Hynda, Kotsiubynskyi and Senkovskyi1968), revised by Walaszczyk et al. (Reference Walaszczyk, Cobban and Odin2002a) and Machalski & Malchyk (Reference Machalski and Malchyk2016), are of particular importance for the stratigraphy of the Nahoryany section (see also Walaszczyk et al., Reference Walaszczyk, Dubicka, Olszewska-Nejbert and Remin2016). These data indicate that the Nahoryany section encompassed two inoceramid zones, namely the upper part of the lower Maastrichtian Endocostea typica Zone and the lower part of the lower/lowermost upper Maastrichtian Trochoceramus radiosus Zone. The inoceramid-based stratigraphy is confirmed by analysis of giant scaphitid taxa associated, on the basis of a comparison with successions recorded from Poland (see Introduction). These suggest that the section at Nahoryany comprised the lower portion of the E. typica Zone as well (Machalski & Malchyk, Reference Machalski and Malchyk2016). The heterogeneous stratigraphical nature of the Nahoryany section was already postulated by Siemiradzki (Reference Siemiradzki1905), who suggested that what he regarded as ‘Scaphites constrictus and tenuistriatus’ occurred only in the upper levels of the ‘Nahoryany opoka’ (Siemiradzki, Reference Siemiradzki1905, pp. 488–489).
The most recent revision of belemnites from Nahoryany brought results that were consistent with the inoceramid and ammonite data. Christensen (Reference Christensen1987) identified Belemnella (Pachybelemnella) inflata (Arkhangelsky, Reference Arkhangelsky1912) in the Nahoryany collections. This species occurs in the Belemnella lanceolata and Belemnella pseudobtusa Zones as distinguished by Schulz (Reference Schulz1979) in the Kronsmoor section (north-west Germany). The currently accepted definition of the base of the Maastrichtian (Odin & Lamaurelle, Reference Odin and Lamaurelle2001) coincides with the base of the Belemnella pseudobtusa Zone sensu Schulz at Kronsmoor (Niebuhr et al., Reference Niebuhr, Hampton, Gallagher and Remin2011, fig. 6). This is the base of the Belemnella obtusa Zone (sensu Niebuhr et al., Reference Niebuhr, Hampton, Gallagher and Remin2011; see also Wilmsen et al., Reference Wilmsen, Engelke, Linnert, Mutterlose and Niebuhr2018), which in the Middle Vistula River section (central Poland) corresponds to the upper portion of the uppermost Campanian ‘I’. redbirdensis Zone and to the lower Maastrichtian E. typica Zone in the inoceramid subdivision (Remin, Reference Remin2012, fig. 2; see also Walaszczyk, Reference Walaszczyk2004). Indeed, markers of the latter zone are well represented in the Nahoryany assemblage (Walaszczyk et al., Reference Walaszczyk, Cobban and Odin2002a; Machalski & Malchyk, Reference Machalski and Malchyk2016).
Material and methods
The present study is based on 18 specimens, which are housed in State Museum of Natural History of the National Academy of Sciences of Ukraine (SMNH-PZ-K) at Lviv (Ukraine), as well as nine specimens at GeoSphere Austria (GSA), formerly Geologische Bundesanstalt, Vienna, Austria. Some of the specimens from the GSA collections were illustrated by Favre (Reference Favre1869) and Kennedy & Summesberger (Reference Kennedy and Summesberger1987). The SMNH specimens studied have never been published, as far as I can tell.
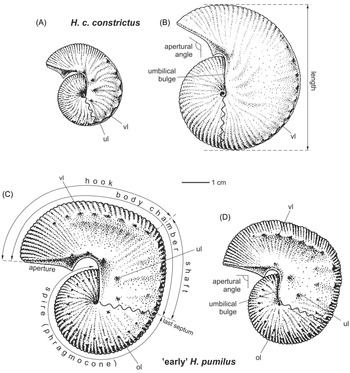
The terminology of the scaphitid conch (Fig. 2A–D) essentially follows that used by Machalski (Reference Machalski2005, Reference Machalski2021) and Machalski & Odin (Reference Machalski, Odin and Odin2001). The dimorphism of each species, presumably of a sexual nature, is accepted. The dimorphs are referred to as macroconchs (M) and microconchs (m) (Fig. 2). Macroconchs typically attain much larger sizes at maturity, have high-whorled body chambers and commonly bear an umbilical bulge (swell) on the dorsal wall of the shaft. Such shells are traditionally interpreted as females. The microconchs, are typically smaller (but not always, as seen by the giant specimen illustrated in Fig. 2C). They are characterised by low-whorled body chambers and concave umbilical walls lacking a bulge, and are regarded as males (e.g., Makowski, Reference Makowski1962; Cobban, Reference Cobban1969; Kennedy, Reference Kennedy1986; Machalski, Reference Machalski1996, Reference Machalski2005, Reference Machalski2021; Landman et al., Reference Landman, Cobban and Larson2012; Klug et al., Reference Klug, Zatoń, Parent, Hostettler, Tajika, Kug, Korn, De Baets, Kruta and Mapes2015). In view of the fact that the material has been largely distorted during diagenesis, only the length of the shell (= maximum diameter in Machalski, Reference Machalski2005, Reference Machalski2021) is measured for complete individuals.

Figure 2. Morphological terms used to describe scaphitid conchs. A, B. Hoploscaphites constrictus. A – microconch; B – macroconch (based on several specimens from the lower upper Maastrichtian Calcaire à Baculites of the Contentin Peninsula, illustrated by Kennedy, Reference Kennedy1986). C, D. Hoploscaphites pumilus. C – giant microconch, characterised by the low height of the shaft whorl; D – macroconch. Specimens C and D are based on specimens from the upper Campanian Saratoga Chalk, illustrated by Kennedy & Cobban, Reference Kennedy and Cobban1993, fig. 17/17–19 for C and fig. 16/22–26 for D. vl – ventrolateral tubercles, ul – umbilical tubercles, ol – outer lateral tubercles.
As is typical of remains of ectocochleate cephalopods in the porous carbonate facies of the European Upper Cretaceous (Malchyk et al., Reference Malchyk, Machalski, Waksmundzki and Duda2017; Janiszewska et al., Reference Janiszewska, Mazur, Machalski and Stolarski2018; Machalski, Reference Machalski2021), all specimens studied are preserved as moulds (steinkerns). More specifically, the Nahoryany specimens are composite moulds with the external ornament superimposed on the internal mould following dissolution of the aragonitic shell and subsequent compaction. Specimens illustrated in Figure 3 were coated with ammonium chloride prior to photography. Line drawings of specimens are largely based on restored (retrodeformed, where needed) individuals from the published collections.

Figure 3. Hoploscaphites pumilus from the lower Maastrichtian of Nahoryany. A, B. GSA2006/086/0002 (m); A – lateral view. B – ventral view. C. GSA1869/006/0011/01 (m), lateral view. D, E. GSA2006/086/0007 (m); D – lateral view, E – ventral view. F–H. GSA2006/086/0001 (M); F, H – lateral view, G – ventral view. I, J. GSA2006/086/0006 (M), I – lateral view, J – ventral view. K–N. GSA1869/006/0011/03 (m); K – ventral view of hook, L, M – lateral views, N – ventral view of shaft. O–R. GSA1869/006/0011/04 (M); O – ventral view of hook, P, Q – lateral views, R – ventral view of shaft. S. GSA2006/086/0008 (m), lateral view. T–V. GSA1869/006/0011/02 (M); T, V – lateral views, U – ventral view of shaft. m – microconch; M – macroconch.
Taxonomy
Three morphologically different groups of specimens are discernible in the material studied. These are assigned here to Hoploscaphites pumilus, H. constrictus and H. sp.
Hoploscaphites pumilus (Stephenson, Reference Stephenson1941)
Description
Nine specimens from GSA collections are assigned here (Fig. 3A–V). Mature microconchs (m), five in total, are identified on the basis of the low whorl of the shaft and the straight or concave umbilical wall (Fig. 3A–E, K–N, S). A distinct gap between the hook and spire is seen in some individuals (Fig. 3L). Some are compressed with flat flanks (Fig. 3A–C), while others are notably depressed and robust in appearance (Fig. 3K–N). The robust specimens are typified by sharply defined ventrolateral and umbilical shoulders and a trapezoidal cross-section of the shaft, resulting from a marked concavity of the flank at the shaft-hook transition. These specimens have also more inflated venters on the hook than on the shaft (compare Fig. 3K,N). Regular and dense ribbing is present over the entire venter. The best-preserved, relatively undistorted microconch (Fig. 3C) is 31 mm long, with an apertural angle close to normal. This is the original of Favre (Reference Favre1869, pl. 5, fig. 1), reillustrated by Kennedy & Summesberger (Reference Kennedy and Summesberger1987, pl. 6, figs. 16–18). Another, much more distorted microconch (Fig. 3K–N) was illustrated by Favre (Reference Favre1869, pl. 5, fig. 4) and Kennedy & Summesberger (Reference Kennedy and Summesberger1987, pl. 6, figs. 19–21).
Mature macroconchs (M), four in total, are identified based on the high whorl of the body chamber (Fig. 3F–J, O–R, T–V). The best-preserved one (Fig. 3T–V) is the original of Favre (Reference Favre1869, pl. 5, fig. 2), reillustrated by Kennedy & Summesberger (Reference Kennedy and Summesberger1987, pl. 6, figs. 13–15). It is a compressed individual, 51 mm long, with a distinct umbilical bulge, flat flanks, rather close contact of the spire and hook, an apertural angle close to normal, and abundant vetrolateral tubercles ranging up to the aperture. Although fragmentary and distorted, a much smaller macroconch (Fig. 3F–H) seems to be similar in proportions and morphology. In contrast, other specimens identified here as macroconchs by virtue of their high-whorled body chambers (e.g., Fig. 3O–R) are strongly depressed, with concave flanks of the shaft and a trapezoidal cross-section at the shaft-hook transition, sharing these characters with the robust microconchs. This is best seen when comparing the microconch in Figure 3K–N and the macroconch in Figure 3O–R. The latter reveals another distinctive feature, namely a break in the pattern of ventrolateral tuberculation corresponding to a change in the geometry of the final whorl. The ventrolateral tubercles are rather distant on the older part of the shaft and more closely spaced over the younger portion of the shaft and hook. This change seems to be associated with an abrupt change in whorl convexity, resulting in the ‘hump-backed’ outline of the specimen (Fig. 3O–R).
Discussion
The material described above matches that of Hoploscaphites pumilus described from the upper Campanian Nacatoch Sand of Navarro County (Texas), Saratoga Chalk of Hempstead County (Arkansas) and Navesink Formation at Atlantic Highlands (New Jersey), all in the USA (Stephenson, Reference Stephenson1941; Cobban, Reference Cobban1974; Kennedy & Cobban, Reference Kennedy and Cobban1993) and that from the upper Campanian and lower Maastrichtian of the D’Avezac and Les Vignes units as exposed at Tercis les Bains, France (Machalski & Odin, Reference Machalski, Odin and Odin2001; Odin et al., Reference Odin, Courville, Machalski, Cobban and Odin2001). There is also a record of ‘Scaphites aff. pumilus Stephenson’ from the upper Campanian of Tajikistan, Middle Asia (Atabekyan & Khakimov, Reference Atabekyan and Khakimov1976, pl. 8, figs. 1, 2), which remains to be verified.
The Saratoga Chalk material of H. pumilus is the most representative in view of its good preservation and abundance. The Saratoga specimens are characterised by a distinctive, rather robust ‘hump-backed’ shell, trapezoidal cross-section of the shaft, an apertural angle close to normal and the occasional presence of an additional row of tubercles next to the ventrolateral ones (outer lateral tubercles; Fig. 2). One of the Saratoga specimens interpreted as a macroconch by Kennedy & Cobban (Reference Kennedy and Cobban1993, fig. 17/17–19) has been reinterpreted by Machalski & Odin (Reference Machalski, Odin and Odin2001) as a giant microconch, marking a notable reversal in the microconch vs macroconch size-relationship typical for scaphitids and other ammonites (Fig. 2). In contrast, the Tercis les Bains material is more variable, comprising ‘Saratoga-type’ specimens in the uppermost Campanian part of the succession (Odin et al., Reference Odin, Courville, Machalski, Cobban and Odin2001, pl. 1, figs. 9–10, 11–13) and a mixture of ‘Saratoga-like’ specimens (e.g., Machalski & Odin, Reference Machalski, Odin and Odin2001, pl. 1, figs. 3, 4, 30–31, 36) with more slender, less ‘hump-backed’ individuals in the lower Maastrichtian portion of the section (Machalski & Odin, Reference Machalski, Odin and Odin2001, pl. 1, figs. 15–16, 25, 26), which is in the Endocostea typica Zone (Walaszczyk et al., Reference Walaszczyk, Odin and Dhondt2002b).
Some of the robust Nahoryany specimens, especially a microconch illustrated in Figure 3K–N, and a macroconch in Figure 3O–R, have close counterparts in the Saratoga material of H. pumilus illustrated by Kennedy & Cobban (Reference Kennedy and Cobban1993, figs. 16–17). The only significant differences are that (1) none of the Nahoryany specimens reveal an additional row of tubercles and (2) the Nahoryany microconchs never exceed in size the associated macroconchs.
The distorted, fairly large microconch specimen figured by Kner (Reference Kner1852, pl. 15, fig. 13) under the name of ‘Scaphites constrictus d’Orb. Var.’ seems to represent the ‘Saratoga-like’ morphology as well; it might even possess an additional row of tubercles as pointed out by Machalski & Odin (Reference Machalski, Odin and Odin2001).
Amongst the compressed Nahoryany specimens, the microconch in Figure 3A,B and the macroconch in Figure 3T–V are close in their appearance to several of the Tercis les Bains specimens, as exemplified by a microconch from a level 10 cm above the Campanian–Maastrichtian boundary (Machalski & Odin, Reference Machalski, Odin and Odin2001, pl. 1, figs 15, 16) and a macroconch from a level 50 cm below this boundary (Machalski & Odin, Reference Machalski, Odin and Odin2001, pl. 1, figs. 25, 26).
The compressed specimens from Nahoryany and Tercis les Bains are also close in their morphology to Hoploscaphites constrictus anterior Błaszkiewicz, Reference Błaszkiewicz1980 from the lower Maastrichtian of the Miechów Upland, southern Poland; this is from the upper part of the inoceramid Endocostea typica Zone (Agata Jurkowska, personal communication, 2018). Indeed, Błaszkiewicz (Reference Błaszkiewicz1980) placed specimens from Nahoryany illustrated by Favre (Reference Favre1869) in the synonymy of H. c. anterior, albeit with a query. Błaszkiewicz (Reference Błaszkiewicz1980) differentiated this subspecies from the nominate one, among other things, by its smaller apertural angle and less close contact of the body chamber and spire. It must be noted, however, that Błaszkiewicz (Reference Błaszkiewicz1980) illustrated only two well-preserved macroconch specimens of his subspecies, which differ in several ways from each other. The holotype (Błaszkiewicz, Reference Błaszkiewicz1980, pl. 18, figs. 4–6), from Polichno near Pińczów, has an open apertural angle, unlike the Saratoga specimens of H. pumilus, but similar to H. constrictus material from the lower upper Maastrichtian Calcaire à Baculites of Cotentin (Kennedy, Reference Kennedy1986). Indeed, the latter author regarded H. c. anterior as a synonym of H. c. constrictus. Another macroconch of H. c. anterior from Śladów (Błaszkiewicz, Reference Błaszkiewicz1980, pl. 18, figs. 9–10) has, however, an apertural angle that is close to normal and a slightly ‘hump-backed’ shell outline, like the Tercis les Bains and Nahoryany macroconchs of H. pumilus discussed above. These characters led Machalski & Odin (Reference Machalski, Odin and Odin2001) and Machalski (Reference Machalski and Bogdanova2019) to postulate that H. constrictus evolved from H. pumilus. However, in opposition to the conclusions reached by Machalski & Odin (Reference Machalski, Odin and Odin2001), H. constrictus anterior is still regarded herein as a doubtful taxon, awaiting revision based on more abundant material from the type locality, which is inaccessible today.
Hoploscaphites constrictus (Sowerby, Reference Sowerby1817)
Description
Six individuals from the SMNH collections are assigned here (Fig. 4A–G), two of these being undoubted macroconchs (Fig. 4A–C) and another one probably a macroconch as well (Fig. 4D); the remaining specimens are microconchs in view of their low whorl of the shaft (Fig. 4E–G). The specimens are rather small when compared to Nahoryany material assigned to H. pumilus.

Figure 4. Scaphitid ammonites, H. constrictus (A–G) and H. sp. (H–N), from the lower Maastrichtian of Nahoryany. A. SMNH-PZ-K-9716/2 (M), lateral view. B, C. SMNH-PZ-K-9716/5 (M), lateral views. D. SMNH-PZ-K-9718/2 (M?), lateral view. E. SMNH-PZ-K-9716/4 (m), lateral view. F. SMNH-PZ-K-9716/1 (m), lateral view. G. SMNH-PZ-K-9716/3 (m), lateral view. H–J. SMNH-PZ-K-9716/3; H – ventral view of spire, I – lateral view, J – ventral view of hook. K. SMNH-PZ-K-9718/3 (M), lateral view, L–N. SMNH-PZ-K-9720 (M); L – ventral view of hook, M – lateral view, N – adapertural view. m – microconch; M – macroconch.
The best-preserved macroconch is shown in Figure 4A. It is 30 mm long, almost circular in lateral view and has a distinct umbilical swelling and flat flanks covered with delicate ribs; barely discernible traces of two or three ventrolateral tubercles are seen in the middle portion of the shaft. Delicate flexuous riblets cover the hook, and the apertural angle is open (c. 120˚). Another specimen (Fig. 4B,C) is closely similar in morphology and size (32 mm in length), with an open apertural angle as well. The specimen bears more accentuated ventrolateral tubercles than the preceding one; umbilical bullae are also visible. The ornament of the incomplete specimen in Figure 4D is stronger.
The best-preserved microconch (Fig. 4F) is of comparable size to the macroconchs, sharing with them faint traces of tuberculation in the middle of the shaft and an open apertural angle. The other two microconchs (Fig. 4E,G), both distorted and fragmentary, are very finely ribbed with no tubercles at all; poor preservation precludes interpretation of the apertural angle.
Discussion
All specimens described above share rather small dimensions, delicate ornament with feeble or missing ventrolateral tuberculation and an open umbilical angle, where this can be seen. They were placed in the same box in the SMNH collections and labelled ‘Scaphites tenuistriatus’. Indeed, the fine, occasionally thread-like ribbing of these individuals recalls that of Hoploscaphites tenuistriatus (Kner, Reference Kner1848). This species was recorded from Nahoryany by Kennedy & Summesberger (Reference Kennedy and Summesberger1987). However, their specimen actually came from the younger chalk succession of the ‘Lvivskaja svita’, representing the conventional lower upper Maastrichtian Belemnitella junior Zone (Machalski & Odin, Reference Machalski, Odin and Odin2001; Machalski, Reference Machalski2005).
The specimens discussed above match well some of the specimens of Hoploscaphites constrictus from the stratum typicum of this species, namely the lower upper Maastrichtian Calcaire à Baculites of Cotentin (Kennedy, Reference Kennedy1986; Walaszczyk & Kennedy, Reference Walaszczyk and Kennedy2011). The French material shows a remarkable variability in size and style of ornamentation of both dimorphs (Kennedy, Reference Kennedy1986), and in contrast to the Nahoryany material is represented by internal rather than composite moulds, but all these specimens share an open apertural angle and relatively short shaft when compared to H. pumilus, especially in macroconchs. The Nahoryany specimens match particularly well the suite of small and finely ornamented Cotentin specimens illustrated by Kennedy (Reference Kennedy1986, pl. 14, figs. 1–9). It should be noted that amongst the specimens figured in Kennedy (Reference Kennedy1986) only that in his plate 14, figs. 1–4 is an undoubted microconch, the remaining ones better interpreted as very small macroconchs, as judged by the presence of an umbilical bulge.
Two of the specimens from the GSA Nahoryany collections illustrated by Kennedy & Summesberger (Reference Kennedy and Summesberger1987, pl. 6, figs. 6–7 and 8–9) as H. constrictus, both undoubted microconchs (not localised by myself in the collections), share the characters of the SMNH specimens discussed herein. All in all, it seems that only small (with length of around 30 mm) and finely ornamented representatives of this species occur at Nahoryany. For comparison, the type material of the species from Cotentin ranges from 22 to 68 mm in shell length (Kennedy, Reference Kennedy1986).
Hoploscaphites sp.
Description
Depressed macroconch individuals from SMNH collections, three in total, are assigned here (Fig. 4H–J, K, L–N). The stout specimen illustrated in Figure 4L–N is 40 mm long, typified by strong flank ribs, prominently swollen inner flanks and ventrolateral tubercles ranging all around the ventrally exposed portion of the adult shell, including particularly strong tubercles on the entire exposed phragmocone (spire). The same extent of tuberculation is visible in the specimen shown in Figure 4H–J. The specimen illustrated in Figure 4L–N is relatively undistorted and has an apertural angle close to normal. The specimen in Figure 4H–J is heavily distorted; incidentally it shows a great similarity to the (lost) specimen illustrated by Kner (Reference Kner1848, pl. 1, fig. 4) as ‘Scaphites compressus d’Orbigny’, which is a specific name coined for a Coniacian scaphitid (see Kennedy, Reference Kennedy1984a).
Discussion
The two macroconchs (Fig. 4H–J, L–N) are utterly different from what is referred to here as H. pumilus and H. constrictus. Instead, they share their strong ‘all-around’ ventrolateral tuberculation with some early Maastrichtian North American scaphitids. These are Hoploscaphites sargklofak Landman, et al., Reference Landman, Kennedy and Larson2015, H. macer Landman et al., Reference Landman, Kennedy, Larson, Grier, Grier and Linn2019 and H. criptonodosus (Riccardi, Reference Riccardi1983). However, the similarity is probably of a superficial nature, because the undeformed Nahoryany specimen (Fig. 4L–N) differs from each of the above-mentioned North American species in having an apertural angle that is close to normal. Specimen in Figure 4K lacks the spire but most probably belongs to this group too.
Discussion on the origin and range of Hoploscaphites constrictus
The subject of the present paper provides an opportunity to discuss the evolutionary origin and stratigraphical range of Hoploscaphites constrictus. As pointed out in Introduction, this widespread and common European scaphitid ammonite was considered as an informal ‘guide fossil’ for the European Maastrichtian as its stratigraphical extent was regarded to be (almost) equal to the range of the Maastrichtian Stage, in the old definition. In order to test this view, an overview of the stratigraphical ranges of the first appearance datum (FAD) of H. constrictus is presented below. The recently elaborated inoceramid stratigraphy of the Maastrichtian Stage (Walaszczyk et al., Reference Walaszczyk, Cobban and Odin2002a, Reference Walaszczyk, Odin and Dhondt2002b; Walaszczyk, Reference Walaszczyk2004; Walaszczyk et al., Reference Walaszczyk, Jagt and Keutgen2010, Reference Walaszczyk, Dubicka, Olszewska-Nejbert and Remin2016; Walaszczyk & Kennedy, Reference Walaszczyk and Kennedy2011; Machalski & Malchyk, Reference Machalski and Malchyk2016; Jagt & Jagt-Yazykova, Reference Jagt and Jagt-Yazykova2018) is accepted as a convenient basis for calibration of scaphitid ranges (Fig. 5).

Figure 5. Succession of temporal subspecies, each represented by a dimorphic pair in their presumed life positions, within the main Hoploscaphites constrictus lineage in the European Maastrichtian. Restored views of successive subspecies are based on Kennedy & Cobban (Reference Kennedy and Cobban1993) for ‘early’ H. pumilus, Machalski & Odin (Reference Machalski, Odin and Odin2001) for ‘late’ H. pumilus, Błaszkiewicz (Reference Błaszkiewicz1980) for H. c. anterior, Kennedy (Reference Kennedy1986) for H. c. constrictus, and Machalski (Reference Machalski2005) for H. c. lvivensis, H. c. crassus and H. c. johnjagti. The scaphitid succession is presented against the inoceramid-based subdivision of the Maastrichtian stage, modified from Landman et al., (Reference Landman, Machalski and Whalen2021, fig. 2, originally compiled from data provided by Abdel-Gawad, Reference Abdel-Gawad1986; Walaszczyk et al., Reference Walaszczyk, Dubicka, Olszewska-Nejbert and Remin2016, fig. 3; Machalski & Malchyk, Reference Machalski and Malchyk2016, fig. 4, Gale et al., Reference Gale, Mutterlose, Batenburg, Gradstein, Agteberg, Ogg, Petrizzio, Gradstein, Ogg, Schmitz and Ogg2020, fig. 27.9, and by Ireneusz Walaszczyk, personal communication, 2021). Scaphitid ranges are based on the papers quoted above; the precise positioning of H. c. anterior is based on speculative extrapolation of the trend in addition to the data presented by Błaszkiewicz (Reference Błaszkiewicz1980). H., Hoploscaphites; c., constrictus; E., Endocostea; Tr., Trochoceramus, I., ‘Inoceramus’; S., Spyridoceramus; T., Tenuipteria; m – microconch, M – macroconch.
The Nahoryany section is not very conclusive with respect to the stratigraphical ranges of the species involved. Based on inoceramid and ammonite data, the section encompassed two zones, namely the lower Maastrichtian Endocostea typica Zone and the lower part of the lower/lowermost upper Maastrichtian Trochoceramus radiosus Zone (Walaszczyk et al., Reference Walaszczyk, Cobban and Odin2002a; Machalski & Malchyk, Reference Machalski and Malchyk2016; Walaszczyk et al., Reference Walaszczyk, Dubicka, Olszewska-Nejbert and Remin2016). The exact provenance level of the studied scaphitid specimens is not available on museum labels. Therefore, it is not known whether they occurred together or formed a succession. It is only certain that H. constrictus appeared in one of these two inoceramid zones. For comparison, the stratum typicum of the species, the Calcaire à Baculites of Cotentin, represents the lower upper Maastrichtian ‘Inoceramus’ ianjoanensis Zone (Walaszczyk & Kennedy, Reference Walaszczyk and Kennedy2011).
At Tercis les Bains, there are no unequivocal records of H. constrictus from correlatives of the E. typica Zone (Machalski & Odin, Reference Machalski, Odin and Odin2001), the base of which is a good proxy for the Campanian-Maastrichtian boundary (Walaszczyk et al., Reference Walaszczyk, Odin and Dhondt2002b).
At Kronsmoor (north-west Germany), most of the specimens identified as H. constrictus by Niebuhr (Reference Niebuhr2003, pl. 7, figs. 2–4) are regarded here as equivocal in view of their poor preservation and lack of diagnostic characteristics. These doubtful individuals include two specimens (Niebuhr, Reference Niebuhr2003, pl. 7, figs 2, 5) found in the interval c. 3.5–4 m above the base of the conventional Belemnella lanceolata Zone, regarded formerly as the base of the Maastrichtian Stage in Europe. The only indisputable specimen of H. constrictus from Kronsmoor is a single macroconch from the Belemnella obtusa Zone (Niebuhr, Reference Niebuhr2003, pl. 7, fig. 4), which is equivalent to the E. typica Zone (Remin, Reference Remin2012).
At Krampen near Neuberg (Steiermark, Austria), the undoubtedly early Maastrichtian ammonite fauna contains H. constrictus, but its precise dating is equivoval (Kennedy & Summesberger, Reference Kennedy and Summesberger1986, see also Machalski et al., Reference Machalski, Jagt, Landman and Motchurova-Dekova2007). Moreover, it has not yet been calibrated against the inoceramid zonation. Some of the specimens regarded as H. constrictus by Kennedy & Summesberger (Reference Kennedy and Summesberger1986, pl. 16, figs. 8, 9 and 13) have subsequently been reinterpreted by Machalski et al. (Reference Machalski, Jagt, Landman and Motchurova-Dekova2007) as an unknown Hoploscaphites species with North American affinities. However, the remaining specimens illustrated as H. constrictus by Kennedy & Summesberger (Reference Kennedy and Summesberger1986, pl. 16, figs. 1–5, 13), all minute macroconchs, belong to this species without doubt.
In Poland, the heterogenic and controversial type material of H. constrictus anterior (discussed in Taxonomy) comes from the upper portion of the E. typica Zone.
At Volsk (Saratov region, Russian Federation), a rich scaphitid fauna has been documented (Selt’ser & Ivanov, Reference Selt’ser and Ivanov2010; Selt’ser & Machalski, Reference Selt’ser, Machalski and Bogdanova2019). This succession includes H. pumilus and H. constrictus. The first unequivocal records of the latter species stem from equivalents of the upper portion of the E. typica Zone (Vladimir Selt’ser, personal communication, 2018; Machalski, Reference Machalski and Bogdanova2019).
In summary, the evidence available to date points to the FAD of Hoploscaphites constrictus somewhere in the E. typica Zone, most probably in its upper part. This zone is the lowermost Maastrichtian inoceramid zone in Europe in the currently accepted subdivision (e.g., Walaszczyk et al., Reference Walaszczyk, Cobban and Odin2002a, Reference Walaszczyk, Odin and Dhondt2002b). There is no evidence that J. Sowerby’s species appeared close to the formerly accepted lower boundary of the Maastrichtian, which is near the base of conventional Belemnella lanceolata Zone.
A view that Hoploscaphites constrictus represents an example of evolutionary stasis (Kennedy, Reference Kennedy1989) cannot be upheld in face of subsequent works. Machalski (Reference Machalski2005, Reference Machalski, Jagt and Jagt-Yazykova2012b) divided the upper part of the H. constrictus lineage into a succession of temporal subspecies (chronosubspecies) of biostratigraphical significance based on rich ‘populations’ from various locations and levels in central Europe. These range from H. constrictus constrictus as understood based on the Cotentin material to H. c. johnjagti, which even passed across the Cretaceous–Paleogene (K–Pg) boundary (Fig. 5). As far as the lower part of the succession in Figure 5 is concerned, it is admittedly speculative in view of uncertainties in precise positioning of the Nahoryany specimens of H. pumilus and H. constrictus and the controversial status of H. constrictus anterior from Poland. What is referred to as ‘early’ H. pumilus in Figure 5 refers to the robust and distinctively ‘hump-backed’ specimens known from the upper Campanian of the Saratoga Chalk and the upper Campanian and lowermost Maastrichtian of the Tercis les Bains section, and from the lower Maastrichtian of Nahoryany. What is shown as ‘late’ H. pumilus in Figure 5 refers to more slender and less ‘hump-backed’ early Maastrichtian specimens identified at Tercis les Bains and Nahoryany. These ‘late’ forms are morphologically similar to H. constrictus, and considered along H. constrictus anterior, which seems to be even more closely similar to H. constrictus constrictus, suggesting an evolutionary transition from H. pumilus to H. constrictus as proposed by Machalski & Odin (Reference Machalski, Odin and Odin2001) and Machalski (Reference Machalski and Bogdanova2019). However, the chronological succession from ‘late’ H. pumilus through H. constrictus anterior to H. c. constrictus presented in Figure 5 is based on arbitrary extrapolation of morphologies and should be tested in future, ideally based on analysis of stratigraphically better-constrained ‘populations’.
If the interpretation of the evolutionary transition from H. pumilus to H. constrictus is correct, some considerations are possible about its significance in terms of functional morphology and mode of life of these cephalopods. According to Landman et al. (Reference Landman, Kennedy, Cobban and Larson2010) and Landman et al. (Reference Landman, Cobban and Larson2012), scaphitid species with long shaft and hook separated from the spire were poorly designed for active horizontal swimming, relying more on passive vertical scanning of the water column. The long shaft meant that the centres of buoyancy and mass of the adult shell were located relatively far from each other, enhancing the hydrostatic stability of the animal, better suitable for a life as a rather passive ‘vertical scanner’ than an active ‘horizontal swimmer’. The conchs of H. pumilus, especially those of the ‘early’ forms of the species (Fig. 5), were robust, heavily ornamented, with the hook well separated from the spire (especially in microconchs), thus representing the category of ‘poor horizontal swimmers’. In contrast, more slender and less heavily ornamented scaphitids with short shafts and closer contacts of the hook and phragmocone may be interpreted as better swimmers, able to do more efficient horizontal movements (Landman et al., Reference Landman, Cobban and Larson2012). The recoiled shells of H. constrictus constrictus and younger subspecies fall into this category (Figs. 2, 5). In summary, the shortening of the body chamber and associated recoiling of the shell during the transition from H. pumilus to H. constrictus enhanced the active horizontal swimming ability, and perhaps also the manoeuvrability of these cephalopods.
As pointed out by Wiedmann (Reference Wiedmann1965), Landman et al. (Reference Landman, Cobban and Larson2012), Landman et al. (Reference Landman, Plint and Walaszczyk2017) and Landman et al. (Reference Landman, Machalski and Whalen2021), many lineages of Late Cretaceous scaphitid ammonites showed a tendency to recoil the final body chamber during their evolution. A recoiling trend, for instance, was demonstrated for the lineage of giant European scaphitids (Landman et al., Reference Landman, Machalski and Whalen2021, fig. 2) mentioned in Introduction.
Final remarks
The early Maastrichtian scaphitid fauna from Nahoryany, south of Lviv (western Ukraine), has been described with three species identified: Hoploscaphites pumilus, H. constrictus and H. sp. All specimens studied came from the old museum collections which lack data on their provenance level within the section, so it cannot be determined whether or not they occurred together or formed a succession. Inoceramid data suggest that the section once exposed at Nahoryany encompassed two successive, essentially lower Maastrichtian inoceramid zones. In view of this, and taking into account the better-constrained records of both species from other European sections, it could be assumed that at Nahoryany H. pumilus came from layers older than H. constrictus (see Fig. 5). In this context, it is intriguing that all specimens identified here as H. pumilus come exclusively from the GSA collection, containing specimens collected (probably well) before the publication of Favre’s monograph in 1869. As far as I know, the SMNH collections were largely assembled later on, possibly during the last decades of the nineteenth century. Perhaps, different portions of the section were accessible for fossil collecting at different times.
The ancestor–descendant relationship between H. pumilus and H. constrictus is proposed in the present paper, following Machalski & Odin (Reference Machalski, Odin and Odin2001). The inferred evolutionary transition was towards a less robust, recoiled shell, which in currently accepted interpretations of the scaphitid palaeobiology, would have enhanced the horizontal swimming ability and manoeuvrability of these cephalopods. As mentioned earlier, the recoiling trends have been documented in several lineages of scaphitid ammonites. The conceivable explanations of such trends may involve response to physical factors (e.g., changes in bathymetry) or to biological interactions (e.g., predation, competition), but this is the theme for another study.
Acknowledgements
For assistance and help in various ways during this project, I am greatly indebted to Julia Didenko, Anatol Mamchur and Sofia Bakayeva (State Museum of Natural History of the National Academy of Sciences of Ukraine, Lviv, Ukraine); the late Herbert Summesberger, Alexander Lukeneder, Mathias Harzhauser and Alice Schumacher (Natural History Museum, Vienna, Austria); Holger Gebhardt and Irene Zorn (GeoSphere Austria, formerly Geologische Bundesanstalt, Vienna, Austria); Vladimir B. Selt’ser (Saratov State University, Saratov, Russian Federation); Neil H. Landman (American Museum of Natural History, New York, USA); Agata Jurkowska (AGH University of Science and Technology in Kraków, Poland); Ireneusz Walaszczyk and Bogusław Waksmundzki (Geology Department, University of Warsaw, Poland) and, last but not least, Grażyna Dziewińska, Marian Dziewiński, and Aleksandra Hołda-Michalska (Institute of Paleobiology, Polish Academy of Sciences, Warsaw, Poland). Two anonymous journal reviewers are thanked for their insightful comments.





 Open access
Open access